METODE UZGOJA MIKROORGANIZMA. STUDIJ KULTURNE I BIOKEMIJSKE
SVOJSTVA
Uzgoj, odnosno uzgoj mikroorganizama u laboratoriju, koristi se za proučavanje njihovih svojstava i dobivanje biomase. Bakterije, gljive, aktinomicete, spirohete i neke protozoe uzgajaju se na hranjivim podlogama. Klamidija, rikecija, virusi i neke protozoe mogu se razmnožavati samo u tijelu životinje ili u živim stanicama.
Kulturna svojstva ove vrste mikroorganizama su: 1) uvjeti potrebni za reprodukciju i 2) priroda rasta na hranjivim podlogama. Kulturna svojstva jedna su od karakteristika koje se uzimaju u obzir pri identificiranju (određivanju vrste) mikroorganizama.
Mediji za kulturu
Mediji za kulturu moraju ispunjavati određene zahtjeve. Moraju sadržavati sve hranjive tvari potrebne za reprodukciju ove vrste mikroba. Neki patogeni mikroorganizmi rastu na jednostavnim hranjivim medijima, dok je drugima za njihovu reprodukciju potreban dodatak krvi, krvnog seruma i vitamina.
U mediju za uzgoj moraju se stvoriti određeni uvjeti dodavanjem otopine natrijevog klorida ili pufera. Za većinu bakterija povoljan je hranjivi medij koji sadrži 0,5% natrijevog klorida. Reakcija hranjivog medija, povoljnog za većinu patogenih bakterija, blago je alkalna, što odgovara pH = 7,2-7,4. Vibrio cholerae raste pri pH = 7,8-8,5, gljive-pri pH = 5-5,5. Mediji za kulturu trebaju biti vlažni, odnosno sadržavati dovoljnu količinu vode, biti što transparentniji i sterilni, odnosno prije sjetve ne smiju sadržavati mikrobe.
Po sastavu i podrijetlu hranjivi su mediji prirodni, umjetni i sintetički. Mediji prirodne kulture prirodni su proizvodi poput krumpira i drugog povrća. Umjetni hranjivi mediji pripremaju se prema specifičnoj recepturi od proizvoda s dodatkom organskih i anorganskih spojeva. Sintetički mediji sadrže određene kemijske spojeve u poznatim koncentracijama.
Po konzistenciji, hranjivi mediji su tekući, polutekući, gusti. Agar-agar-polisaharid izoliran iz algi obično se koristi kao brtvilo. Agar-agar mikroorganizmi ne koriste kao hranjivo; on stvara gel u vodi koji se topi na 100 ° C i učvršćuje na 45 ° C.
Za dobivanje gustog hranjivog medija dodaje se agar-agar u koncentraciji od 1,5-2%, za polutekući-0,5%.
Prema namjeni, kulturološki se mediji mogu podijeliti na obične (jednostavne), posebne, izborne, diferencijalnu dijagnostiku.
Uobičajeni (jednostavni) hranjivi mediji koriste se za uzgoj većine mikroorganizama, to je mezopatamijska juha (MPB), mezopatamijski agar (MPA).
Za uzgoj mikroorganizama koji ne rastu na jednostavnim podlogama koriste se posebni hranjivi mediji. Na primjer, krvni agar i šećerna juha za streptokoke, serumski agar za meningokok i gonokok.
Mediji za izbor kulture koriste se za izolaciju jedne vrste iz mješavine različitih bakterija. Ova vrsta bakterija raste u ovoj okolini brže i bolje od drugih, nadmašujući ih u svom rastu; rast drugih bakterija je odgođen na ovom mediju. Na primjer, koagulirani serum za difterijski bacil, alkalna peptonska voda za koleru vibrio, žučna juha za tifusni bacil, slana podloga za stafilokok.
Hranjivi mediji za diferencijalnu dijagnostiku koriste se za razlikovanje nekih vrsta bakterija od drugih po njihovom enzimskom djelovanju (vidi odgovarajući odjeljak).
Uzgoj i izolacija čistih kultura aerobnih bakterija
Za uzgoj mikroorganizama potrebni su određeni uvjeti: temperatura, aerobni ili anaerobni uvjeti.
Temperatura bi trebala biti optimalna za vrstu. Većina patogenih bakterija uspijeva na 37 ° C. Međutim, za neke je vrste optimalna niža temperatura, što je povezano s osobitostima njihove ekologije. Dakle, za bacil kuge, čije su prirodno stanište glodavci tijekom hibernacije, optimalna temperatura je 28 ° C, kao i za leptospiru, za bacil botulizma - 28 ° C -35 ° C.
Osim optimalne temperature, za uzgoj mikroorganizama, ovisno o vrsti, potrebno je aerobno ili anaerobno okruženje.
Za proučavanje morfologije, kulturnih, biokemijskih i drugih svojstava mikroba potrebno je dobiti čistu kulturu. Obično se kultura mikroba naziva njihovo nakupljanje na hranjivom mediju u obliku zamućenosti, rasta pri dnu (stijenke) ili filma na površini tekućeg medija ili kolonija na gustom mediju. Jedna kolonija nastaje od jedne mikrobne stanice. Čista kultura je kultura mikroba iste vrste dobivena iz jedne kolonije. U laboratorijima se za različite studije koriste određeni poznati sojevi mikroba. Soj je čista kultura mikroba dobivena iz određenog izvora, u određeno vrijeme, s poznatim svojstvima. Obično su mikrobni sojevi označeni određenim brojem. Na primjer, soj Staphylococcus aureus 209P koristi se za određivanje aktivnosti penicilina.
Izolacija čistih kultura aeroba obično traje tri dana i provodi se prema sljedećoj shemi:
1. dan - mikroskopija razmaza iz ispitivanog materijala, obojenog (obično po Gramu) - radi prethodnog upoznavanja s mikroflorom, što može biti korisno pri odabiru podloge za kulturu za inokulaciju. Zatim se inokulira materijal na površini smrznutog hranjivog agara radi dobivanja izoliranih kolonija. Prosijavanje se može provesti prema Drygalsky metodi u tri Petrijeve zdjele s hranjivim medijem. Kap materijala nanosi se na prvu šalicu i lopaticom se razmazuje po cijeloj šalici. Zatim istom lopaticom rasporedite preostalu kulturu po njoj na drugu šalicu i na isti način na treću. Najveći broj kolonija izrast će na prvoj ploči, a najmanje na trećoj. Izolirane kolonije rasti će na jednoj od ploča, ovisno o tome koliko je mikrobnih stanica bilo u ispitivanom materijalu.
Isti rezultat može se postići prosijavanjem na jednu šalicu. Da biste to učinili, podijelite šalicu na četiri sektora. Materijal koji se proučava inokulira se bakteriološkom petljom s potezima na prvom sektoru, zatim se, nakon kalcinacije i hlađenja petlje, inokulacija distribuira iz prvog sektora u drugi i na isti način uzastopno u treći i četvrti sektor. Izolirane kolonije nastaju od pojedinačnih mikrobnih stanica nakon svakodnevne inkubacije u termostatu.
2. dan - proučavanje kolonija uzgojenih na pločama, njihov opis. Kolonije mogu biti prozirne, prozirne ili neprozirne, različitih su veličina, zaobljenih pravilnih ili nepravilnih obrisa, konveksnog ili ravnog oblika, glatke ili hrapave površine, glatkih ili valovitih, nazubljenih rubova. Mogu biti bezbojne ili bijele, zlatne, crvene, žute. Na temelju proučavanja ovih karakteristika odrasle kolonije dijele se u skupine. Zatim se iz ispitivane grupe odabere izolirana kolonija, pripremi se bris za mikroskopsko ispitivanje radi provjere homogenosti mikroba u koloniji. Ista se kolonija inokulira u epruvetu s kosim hranjivim agarom.
3. dan - provjera čistoće kulture uzgojene na kosini agara mikroskopijom razmaza. S homogenošću proučavanih bakterija, izolacija čiste kulture može se smatrati potpunom.
Kako bi se identificirale izolirane bakterije, proučavaju se kulturne osobine, odnosno priroda rasta na tekućim i čvrstim hranjivim medijima. Na primjer, streptokoki na šećernoj juhi tvore dno i tjemeni sediment, na krvnom agaru - male, točno određene kolonije; cholera vibrio tvori film na površini alkalne peptonske vode, a prozirne kolonije na alkalnom agaru; bacil kuge na hranjivom agaru tvori kolonije u obliku "čipkastih rupčića" s gustim središtem i tankim valovitim rubovima, a u tekućem hranjivom mediju - film na površini, a zatim - niti koje se iz njega protežu u obliku " stalaktiti ".
Uzgoj i izolacija čistih kultura anaerobnih bakterija
Za uzgoj anaeroba potrebno je smanjiti oksidacijsko-redukcijski potencijal medija, stvoriti anaerobiozu uklanjanjem kisika fizikalnim, kemijskim ili biološkim metodama.
Fizičke metode uključuju:
1) mehaničko uklanjanje zraka pomoću pumpe iz anae-rostata, u koje se stavlja posuda s inokulacijom. Istodobno možete zamijeniti zrak indiferentnim plinom: dušik, vodik, ugljični dioksid.
2) uzgoj u mediju koji sadrži reducirajuće tvari. Kitta-Tarozzi srijeda je šećerna juha s komadićima jetre ili mesa. Glukoza i dijelovi organa imaju sposobnost reduciranja. Medij se na vrh prelije slojem vazelinskog ulja kako bi se blokirao pristup kisiku zraka.
3) Najjednostavnija, ali manje pouzdana metoda je uzgoj duboko u visokom stupcu šećernog agara.
Kemijske metode sastoje se u tome što se posude s usjevima anaeroba stavljaju u hermetički zatvoreni eksikator, gdje se stavljaju kemikalije, na primjer, pirogalol i lužine, čija reakcija teče apsorpcijom kisika.
Biološka metoda temelji se na istovremenom uzgoju anaeroba i aeroba na krutim hranjivim medijima u Petrijevim posudama, hermetički zatvorenim nakon inokulacije. Prvo rastući aerobi apsorbiraju kisik, a zatim počinje rast anaeroba.
Izolacija čiste kulture anaeroba počinje nakupljanjem anaerobnih bakterija inokulacijom na Kitta-Tarozzijev medij. U budućnosti se izolirane kolonije dobivaju na jedan od dva načina:
1) materijal se cijepi miješanjem s otopljenim toplim šećerom u staklenim epruvetama. Nakon što se agar učvrstio, u njegovoj dubini rastu izolirane kolonije koje se uklanjaju rezanjem cijevi i subkulturiraju na Kitt-Tarozzijevom mediju (Weinbergova metoda);
2) inokulacija materijala provodi se na pločama s hranjivim medijem i inkubira u anaerostatu. Izolirane kolonije uzgojene na ploči subkulturirane su na Kitt-Tarozzijevom mediju (Zeisslerova metoda).
Uzgoj drugih mikroorganizama
Uzgoj mikoplazmi
Mikoplazme se uzgajaju na hranjivim medijima nadopunjenim serumom i ugljikohidratima. Budući da mikoplazmama nedostaje stanična stijenka, one rastu samo u izotoničnim ili hipertoničnim okruženjima. Na krutim hranjivim podlogama nekoliko dana stvaraju se vrlo male kolonije, nalik prženim jajima - s ispupčenim središtem i ravnom prozirnom periferijom. Mikoplazme se također mogu uzgajati u kokošjim zametcima ili staničnoj kulturi.
Uzgoj rikecije i klamidije
Rikecije i klamidije obvezni su unutarstanični paraziti. Za njihov uzgoj koriste se stanične kulture, pileći zametci i infekcije životinja.
Uzgoj gljiva
Za uzgoj gljiva koriste se gusti i tekući hranjivi mediji: najčešće Sabouraudov medij, kao i mediji koji sadrže pivsku sladovinu. Gljive rastu sporije od bakterija, formiraju vidljiv rast u roku od nekoliko dana. Temperatura uzgoja niža je od temperature bakterija - 22-30 ° C.
Uzgoj spiroheta i protozoa
Među spirohetama je najlakše uzgojiti leptospiru, kojoj voda pomiješana sa serumom krvi kunića može poslužiti kao hranjivi medij.Borelije i treponeme uzgajaju se u anaerobnim uvjetima na složenijim hranjivim medijima koji sadrže serum, komadiće životinjskog tkiva.
Među protozoama, na hranjivim podlogama uzgajaju se dizenterijska ameba, lamblija, trihomonas, lajšmanija, tripanosom, balantidija, dok se toksoplazma uzgaja u kokošjim zametcima i kulturama tkiva. Metode uzgoja malarijske plazmodije su u razvoju.
Metode proučavanja enzimske aktivnosti (biokemijska svojstva)
U mikrobiološkoj praksi proučavanje enzimske aktivnosti koristi se za identifikaciju mikroorganizama, budući da svaka mikrobna vrsta ima određeni skup enzima.
Za određivanje proteolitičke aktivnosti mikrobi se inokuliraju injekcijom u stupac želatine, a nakon 3-5 dana inkubacije na sobnoj temperaturi primjećuje se karakter ukapljivanja želatine: u obliku lijevka, čavla, čarape ili u obliku prevrnutog božićnog drvca. Proteolitička aktivnost određena je i stvaranjem produkata razgradnje proteina: indola, sumporovodika, amonijaka. Kako bi se odredili, mikroorganizmi se inokuliraju u mesno-peptonsku juhu, a indikatorski papiri stavljaju se između grla epruvete i pamučnog čepa, isključujući njihov kontakt s medijem. Kad nastane indol, papir impregniran zasićenom otopinom oksalne kiseline postaje ružičast; u prisutnosti sumporovodika, papir impregniran olovnim acetatom postaje crn; pri stvaranju amonijaka crveni lakmus papir postaje plav.
Za određivanje saharolitičkih svojstava mikroba koriste se diferencijalno dijagnostički mediji, poput Gissovog medija, Olkenitskog, Endovog, Levinovog, Ploskirevljevog medija.
Mediji Endo, Levin, Ploskirev u Petrijevim posudama koriste se za razlikovanje bakterija crijevne skupine prema njihovoj sposobnosti fermentacije laktoze. Ovi mediji sadrže hranjivi agar, laktozu i indikator koji mijenja boju u kiselom mediju - pokazatelj pH. Ako posijete bakterije koje fermentiraju laktozu, poput E. coli, u takvo okruženje, kiselina nastaje kao posljedica fermentacije laktoze, a indikator će promijeniti boju u kiselom okruženju. Stoga će kolonije Escherichia coli na takvim medijima biti obojene prema boji indikatora: na Endovom i Ploskirevovom mediju - u crvenoj boji, na Levinovoj podlozi - u crnoj i plavoj boji. Kolonije bakterija koje ne fermentiraju laktozu, poput salmonele i dizenterije, bit će bezbojne.
Gissovi mediji (mediji s raznolikim rasponom) pripremaju se na bazi peptonske vode ili polutekućeg mesno-peptonskog agara. Sadrži bilo koji ugljikohidratni ili polihidrični alkohol i indikator. Kad na Gissovom mediju raste mikrob, fermentirajući ovaj supstrat stvaranjem kiseline i plina, medij će promijeniti boju, u polutekućem mediju pojavit će se mjehurići i puknuća u debljini agara, u tekućem mediju - mjehurić plina u staklenom plovku. Kada se supstrat fermentira samo u kiselinu, dolazi samo do promjene boje medija.
Također se koriste i kombinirani mediji koji ne sadrže jedan ugljikohidrat, već dva ili tri, na primjer, medij Olkenitskog. Jedna cijev ovog medija zamjenjuje agar koso i Giss medij laktozom, glukozom i saharozom. Nakon sterilizacije u rastaljenom stanju, medij u epruveti se zakosi tako da se dobije stupac i ukošeni dio. Sjetva se vrši potezom na skošenom dijelu i ubodom u stupac. Prilikom fermentacije laktoze ili saharoze mijenja se boja cijelog medija; kada se fermentira samo glukoza, mijenja se samo boja stupca. Formiranje plina označeno je prisutnošću mjehurića u stupcu agara. Kada mikrobi otpuštaju amonijak, boja medija se ne mijenja. Stvaranje sumporovodika očituje se pocrnjenjem u tablici s agarom
Za ekspresnu metodu za određivanje enzimske aktivnosti bakterija koriste se sustavi mikrotesta i sustav indikatorskog papira (NIB)
Sustav mikrotesta je spremnik od prozirnog polistirena, koji se sastoji od nekoliko stanica. Stanice sadrže osušene hranjive medije s ugljikohidratima i pH pokazateljima. U svaku ćeliju se cijepi suspenzija kulture bakterija određene gustoće. Ulije se slana otopina u kontrolne ćelije. boje
indikator
Sustavi s indikatorskim papirom (NIB) za identifikaciju obitelji enterobacteriaceae su diskovi ili trake od kromatografskog papira, prekrivene zaštitnom folijom koje sadrže određenu podlogu i indikator. Promjenom boje indikatora Za određivanje sumporovodika, disk je postavljen na površinu MPA -e, zasijan injekcijom, što vam omogućuje istodobno određivanje pokretljivosti
U svim epruvetama uzima se u obzir preliminarni rezultat istog dana i konačni rezultat sljedećeg dana.
Aktivnost oksidaze određuje se mljevenjem kulture na indikatorskom papiru, a rezultat se uzima u obzir nakon minute.
Mikroorganizmi (s izuzetkom obligatnih unutarstaničnih parazita - rikecija, klamidija, virusi i protozoe) obično se uzgajaju na umjetnim hranjivim podlogama. Ovisno o prehrambenim potrebama jedne ili druge vrste hranjivih podloga trebale bi sadržavati odgovarajuće polazne tvari potrebne za metabolizam plastike i energije.
Izolacija mikroorganizama iz različitih materijala i proizvodnja njihovih kultura naširoko se koriste u laboratorijskoj praksi za mikrobiološku dijagnostiku zaraznih bolesti, u istraživačkom radu te u mikrobiološkoj proizvodnji cjepiva, antibiotika i drugih biološki aktivnih proizvoda vitalne aktivnosti mikroba.
Uvjeti uzgoja također ovise o svojstvima dotičnih mikroorganizama. Većina patogenih mikroba uzgaja se na hranjivim medijima na 37 ° C 12 dana. Međutim, nekima od njih je potrebno duže vrijeme isporuke. Na primjer, bakterije hripavca - za 2-3 dana, i mycobacterium tuberculosis - za 3-4 tjedna.
Za poticanje procesa rasta i razmnožavanja aerobnih mikroba, kao i za smanjenje vremena njihova uzgoja, koristi se metoda uzgoja pod vodom, koja se sastoji u kontinuiranom prozračivanju i miješanju hranjivog medija. Dubinska metoda našla je široku primjenu u biotehnologiji.
Za uzgoj anaeroba koriste se posebne metode čija je bit uklanjanje zraka ili zamjena inertnim plinovima u zatvorenim termostatima - anaerostatima. Anaerobi se uzgajaju na hranjivim podlogama koje sadrže reducirajuće tvari (glukoza, natrijev formijat itd.) Koje smanjuju redoks potencijal.
U dijagnostičkoj praksi od posebne su važnosti čiste kulture bakterija koje se izoliraju iz ispitivanog materijala uzetog s pacijenta ili objekata iz okoliša. U tu se svrhu koriste umjetni hranjivi mediji, koji se dijele na osnovne, diferencijalno dijagnostičke i izborne medije najrazličitijeg sastava. Odabir hranjivog medija za izolaciju čiste kulture bitan je za bakteriološku dijagnostiku.
U većini slučajeva koriste se čvrste podloge za kulturu, prethodno sipane u Petrijeve zdjele. Ispitni materijal stavlja se na površinu medija u petlju i triturira lopaticom da se dobiju izolirane kolonije izrasle iz jedne stanice. Subkultura izolirane kolonije na kosini agara u epruveti rezultira čistom kulturom.
Za identifikaciju, tj. Utvrđujući generičku i vrstu koje pripada odabranoj kulturi, najčešće se proučavaju fenotipske karakteristike:
a) morfologiju bakterijskih stanica u obojenim mrljama ili nativnim pripravcima;
b) biokemijski znakovi kulture prema sposobnosti fermentacije ugljikohidrata (glukoza, laktoza, saharoza, maltoza, manitol itd.), za stvaranje indola, amonijaka i sumporovodika, koji su produkti proteolitičke aktivnosti bakterija.
Za potpuniju analizu koriste se plinsko-tekućinska kromatografija i druge metode.
Uz bakteriološke metode identifikacije čistih kultura, naširoko se koriste imunološke metode istraživanja koje su usmjerene na proučavanje antigene strukture izolirane kulture. U tu svrhu koriste se serološke reakcije: aglutinacija, imunofluorescentna taloženje, vezanje komplementa, imunološki test enzima, metode radioimunološkog testa itd.
-
Metode izolacije čiste kulture
Kako bi se izolirala čista kultura mikroorganizama, potrebno je međusobno odvojiti brojne bakterije koje se nalaze u materijalu. To se može postići metodama koje se temelje na dva načela -mehanički ibiološki disocijacija bakterija.
Metode izolacije čistih kultura temeljene na mehaničkom principu
Metoda serijskog razrjeđivanja, koju je predložio L. Pasteur, bila je jedna od prvih, koja se koristila za mehaničko odvajanje mikroorganizama. Sastoji se od provođenja serijskog serijskog razrjeđivanja materijala koji sadrži mikrobe u sterilnom uređajutekućinahranjivi medij. Ova je tehnika prilično mukotrpna i nesavršena u radu jer vam ne dopušta kontrolu broja mikrobnih stanica koje ulaze u epruvete tijekom razrjeđivanja.
Nema ovaj nedostatakKochova metoda (metoda razrjeđivanja ploča). R. Koch je koristio krute hranjive podloge na bazi želatine ili agar-agara. Materijal s asocijacijama različitih vrsta bakterija razrijeđen je u nekoliko epruveta otopljenom i blago ohlađenom želatinom čiji se sadržaj kasnije izlio na sterilne staklene ploče. Nakon želiranja medija, uzgojen je na optimalnoj temperaturi. Izolirane kolonije mikroorganizama nastale su u njegovoj debljini, koje se lako mogu prenijeti u svježi hranjivi medij pomoću platinske petlje kako bi se dobila čista kultura bakterija.
Drygalskijeva metodaje naprednija metoda koja se naširoko koristi u svakodnevnoj mikrobiološkoj praksi. Prvo se ispitivani materijal pipetom ili petljom nanese na površinu medija u Petrijevoj zdjelici. Metalnom ili staklenom lopaticom dobro je utrljajte u medij. Posuda se tijekom sjetve drži otvorena i lagano zakreće kako bi se materijal ravnomjerno rasporedio. Bez sterilizacije lopatice, posuđeni materijal provode u drugoj Petrijevoj zdjelici, po potrebi u trećoj. Tek tada se lopatica umoči u otopinu za dezinfekciju ili prži na plamenu plamenika. Na površini medija, u prvom jelu, u pravilu promatramo kontinuirani rast bakterija, u drugom - gusti rast, a u trećem - rast u obliku izoliranih kolonija.
Kolonije Drigalski
Metoda linijske kulturedanas se najčešće koristi u mikrobiološkim laboratorijima. Materijal koji sadrži mikroorganizme sakuplja se bakteriološkom petljom i nanosi na površinu medija za uzgoj blizu ruba posude. Uklonite višak materijala i držite ga paralelnim potezima od ruba do ruba čaše. Nakon jednog dana inkubacije inokulacija na optimalnoj temperaturi, izolirane kolonije mikroba rastu na površini posude.
Metoda moždanog udara
Za dobivanje izoliranih kolonija možete koristiti prekriveni bris koji je korišten za prikupljanje ispitivanog materijala. Malo otvorite Petrijevu zdjelu s hranjivim medijem, umetnite tampon u nju i pažljivo utrljajte materijal u površinu posude, postupno vraćajući tampon i posudu.
Dakle, značajna prednost metoda razrjeđivanja ploča Kocha, Drygalskog i prugaste kulture je ta što stvaraju izolirane kolonije mikroorganizama, koje se, cijepljenjem na drugi hranjivi medij, pretvaraju u čistu kulturu.
Biološke metode za izolaciju čistih kultura
Biološki princip razdvajanja bakterija predviđa namjerno traženje metoda koje uzimaju u obzir brojne karakteristike mikrobnih stanica. Među najčešćim metodama su sljedeće:
1. Prema vrsti disanja. Svi mikroorganizmi prema vrsti disanja podijeljeni su u dvije glavne skupine:aerobni (Corynebacterium diphtheriaeVibrio cholerae itd) ianaerobni (Clostridium tetaniClostridium botulinumClostridium perfringens i tako dalje.)... Ako se materijal iz kojeg treba izolirati anaerobne patogene prethodno zagrije, a zatim uzgaja u anaerobnim uvjetima, tada će te bakterije rasti.
2. Napisaosporulacija. Poznato je da su neki mikrobi (bacili i klostridije) sposobni za plodnost. Među njimaClostridium tetaniClostridium botulinumClostridium perfringensBacillus subtilisBacillus cereus... Sporovi su otporni na djelovanje čimbenika okoliša. Posljedično, ispitni materijal može biti podložan djelovanju toplinskog faktora, a zatim se inokulativno prenijeti u hranjivi medij. Nakon nekog vremena na njemu će rasti upravo one bakterije koje su sposobne za plodnost.
3. Otpornost mikroba na kiseline i lužine. Neke klice(Mycobacterium tuberculosisMycobacterium bovis) zbog osobitosti njihove kemijske strukture otporne su na djelovanje kiselina. Zato se materijal koji ih sadrži, na primjer, ispljuvak kod tuberkuloze, prethodno obrađuje s jednakim volumenom 10% -tne otopine sumporne kiseline, a zatim sije na hranjive podloge. Vanjska flora odumire, a mikobakterije rastu kao rezultat njihove otpornosti na kiseline.
Kolera vibrio(Vibrio cholerae)naprotiv, to je halofilna bakterija, stoga se za stvaranje optimalnih uvjeta rasta sije na podloge koje sadrže lužine (1% alkalne peptonske vode). Već nakon 4-6 sati na površini medija pojavljuju se karakteristični znakovi rasta u obliku nježnog plavkastog filma.
4. Mobilnost bakterija. Neke klice(Proteus vulgaris) imaju tendenciju puzanja i mogu se brzo širiti po površini donekle vlažnog okoliša. Kako bi se izolirali takvi patogeni, oni se inokuliraju u kapljicu kondenzacijske tekućine, koja nastaje hlađenjem nagiba agara. Nakon 16-18 godina proširili su se na cijelu površinu okoliša. Uzmemo li materijal s vrha agara, imat ćemo čistu kulturu patogena.
5. Osjetljivost mikroba na djelovanje kemikalija, antibiotika i drugih antimikrobnih sredstava.Zbog karakteristika metabolizma bakterija, one mogu imati različitu osjetljivost na određene kemijske čimbenike. Poznato je da su stafilokoki, aerobni bacili koji tvore spore, otporni na djelovanje 7,5-10% natrijevog klorida. Zato se za izolaciju ovih patogena koriste izborni hranjivi mediji (agar od žumanjaka-soli, agar beckon-sol) koji sadrže upravo tu tvar. Druge bakterije praktički ne rastu pri ovoj koncentraciji natrijevog klorida.
6. Primjena nekih antibiotika(nistatin) koristi se za inhibiranje rasta gljivica u materijalu koji je s njima jako kontaminiran. Nasuprot tome, dodavanje antibiotika penicilina u medij potiče rast bakterijske flore ako se izoliraju gljivice. Dodavanjem furazolidona u određenim koncentracijama u hranjivi medij stvaraju se selektivni uvjeti za rast korinebakterija i mikrokoka.
7. Sposobnost mikroorganizama da prodru kroz neoštećenu kožu. Neke patogene bakterije(Yersinia pestis) kao posljedica prisutnosti velikog broja enzima agresije, oni mogu prodrijeti kroz netaknutu kožu. U tu se svrhu obriše vuna na tijelu laboratorijske životinje i utrlja se ispitni materijal u ovo područje koje sadrži patogen i veliku količinu mikroflore trećih strana. Nakon nekog vremena životinja se zakolje, a mikrobi se oslobađaju iz krvi ili unutarnjih organa.
8. Osjetljivost laboratorijskih životinja na uzročnike infekcija.Neke su životinje vrlo osjetljive na razne mikroorganizme.
Na primjer, sa bilo kojim putem primjeneStreptococcus pneumoniaebijeli miševi razvijaju generaliziranu pneumokoknu infekciju. Slična se slika opaža kada su zamorci zaraženi uzročnicima tuberkuloze.(Mycobacterium tuberculosis).
U svakodnevnoj praksi bakteriolozi koriste pojmove kao što sunaprezanjeičista kulturamikroorganizmi. Pod sojem se podrazumijevaju mikrobi iste vrste koji su izolirani iz različitih izvora ili iz istog izvora, ali u različito vrijeme. Čista kultura bakterija su mikroorganizmi iste vrste, potomci jedne mikrobne stanice, koji su rasli na (ne) hranjivom mediju.
Izolacija čiste kulture aerobni mikroorganizmi sastoji se od nekoliko faza.
Prvi dan(1. faza istraživanja) patološki materijal se uzima u sterilnu posudu (epruveta, tikvica, boca). Proučavaju ga - izgled, teksturu, boju, miris i druge znakove, pripremaju razmaz, boje i pregledavaju ga pod mikroskopom. U nekim slučajevima (akutna gonoreja, kuga) u ovoj se fazi može postaviti preliminarna dijagnoza, a osim toga moguće je odabrati medij na koji će se materijal cijepiti. Zatim provode bakteriološku petlju (najčešće korištenu), pomoću lopatice - metodom Drygalsky, s vatom od gaze. Šalice su zatvorene, okrenute naopako, potpisane posebnom olovkom i stavljene u termostat na optimalnoj temperaturi (37 ° C) 18-48 sati. Cilj ove faze je dobivanje izoliranih kolonija mikroorganizama.
Međutim, ponekad se radi gomilanja materijala sije na tekuće hranjive podloge.
Na drugi dan(2. faza istraživanja) na površini guste hranjive podloge mikroorganizmi tvore kontinuirani, gusti rast ili izolirane kolonije.Kolonija - To su nakupine bakterija vidljive golim okom na površini ili u debljini hranjivog medija. U pravilu se svaka kolonija tvori od potomaka jedne mikrobne stanice (klonovi), pa je njihov sastav prilično homogen. Značajke rasta bakterija na hranjivim podlogama očitovanje su njihovih kulturnih svojstava.
Ploče se pomno pregledavaju i pregledavaju na izolirane kolonije koje su narasle na površini agara. Obratite pozornost na veličinu, oblik, boju, prirodu rubova i površine kolonija, njihovu konzistenciju i druge značajke. Ako je potrebno, pregledajte kolonije pod povećalom, malim ili velikim povećanjem mikroskopa. Struktura kolonija ispituje se u propuštenom svjetlu pri malom povećanju mikroskopa. Mogu biti hijalinske, zrnaste, nitaste ili vlaknaste, koje karakterizira prisutnost isprepletenih niti u debljini kolonija.
Karakterizacija kolonija važan je dio rada bakteriologa i laboratorijskog pomoćnika, jer mikroorganizmi svake vrste imaju svoje posebne kolonije.
Trećeg dana(Faza 3 istraživanja) proučiti prirodu rasta čiste kulture mikroorganizama i izvršiti njezinu identifikaciju.
Prvo, obraćaju pozornost na osobitosti rasta mikroorganizama na podlozi i prave razmaz, bojeći ga po Gramovoj metodi, kako bi provjerili čistoću kulture. Ako se pod mikroskopom promatraju bakterije iste vrste morfologije, veličine i svojstava tinktora (sposobnost bojenja), zaključuje se da je kultura čista. U nekim slučajevima, već po izgledu i karakteristikama njihovog rasta, može se zaključiti o vrsti izoliranih patogena. Određivanje vrste bakterija prema njihovim morfološkim karakteristikama naziva se morfološka identifikacija.Određivanje vrste patogena prema njihovim kulturnim obilježjima naziva se kulturna identifikacija.
Međutim, ove studije nisu dovoljne za konačan zaključak o vrsti izoliranih mikroba. Stoga proučavaju biokemijska svojstva bakterija. Prilično su raznoliki.
-
Identifikacija bakterija.
Određivanje vrste patogena prema njegovim biokemijskim svojstvima naziva se biokemijska identifikacija.
Kako bi se ustanovile vrste bakterija, često se proučava njihova antigenska struktura, odnosno identificiraju se po antigenim svojstvima. Svaki mikroorganizam sadrži različite antigene tvari. Konkretno, predstavnici obitelji enterobacteriaceae (Yesherichia, Salmoneli, Shigela) sadrže membranski O-antigen, flagelatni H-antigen i kapsularni K-antigen. Po kemijskom sastavu heterogene su, stoga postoje u mnogim varijantama. Mogu se odrediti pomoću specifičnih aglutinoznih seruma. Ova definicija vrste bakterije naziva se serološka identifikacija.
Ponekad se bakterije identificiraju zaražavanjem laboratorijskih životinja čistom kulturom i promatranjem promjena koje patogeni uzrokuju u tijelu (tuberkuloza, botulizam, tetanus, salmoneloza itd.). Ova metoda se naziva identifikacija po biološkim svojstvima... Kao objekti - najčešće se koriste zamorci, bijeli miševi i štakori.
PRILOZI
(tablice i dijagrami)
Fiziologija bakterija
Shema 1. Fiziologija bakterija.
hrana
dah
visina
reprodukcija
raste na hranjivim podlogama
Tablica 1. Opća tablica fiziologije bakterija.
|
№ |
Koncept |
Karakteristično |
|
Prehrana |
Proces stjecanja energije i tvari. |
|
|
Dah |
Skup biokemijskih procesa, uslijed kojih se oslobađa energija potrebna za vitalnu aktivnost mikrobnih stanica. |
|
|
Visina |
Koordinirano razmnožavanje svih staničnih komponenti i struktura, što u konačnici dovodi do povećanja stanične mase |
|
|
Reprodukcija |
Povećanje broja stanica u populaciji |
|
|
Uzgoj na hranjivim podlogama. |
U laboratorijskim uvjetima mikroorganizmi se uzgajaju na hranjivim podlogama koje moraju biti sterilne, prozirne, vlažne, sadržavati određene hranjive tvari (proteine, ugljikohidrate, vitamine, elemente u tragovima itd.), Imati određeni puferski kapacitet, imati odgovarajući pH, redoks potencijal. |
Tablica 1.1 Kemijski sastav i fiziološke funkcije elemenata.
|
№ |
Element kompozicije |
Karakteristike i uloga u staničnoj fiziologiji. |
|
|
Voda |
Glavna komponenta bakterijske stanice, koja čini oko 80% njene mase. U slobodnom je ili vezanom stanju sa strukturnim elementima ćelije. U sporovima se količina vode smanjuje na 18,20%. Voda je otapalo za mnoge tvari i također ima mehaničku ulogu u stvaranju turgora. Tijekom plazmolize - gubitka vode stanicom u hipertoničnoj otopini - protoplazma se odvaja od stanične membrane. Uklanjanje vode iz stanice, sušenje, obustava metaboličkih procesa. Većina mikroorganizama dobro podnosi sušenje. Uz nedostatak vode, mikroorganizmi se ne razmnožavaju. Sušenje u vakuumu iz smrznutog stanja (liofilizacija) zaustavlja reprodukciju i potiče dugoročno očuvanje mikrobnih jedinki. |
||
|
Protein |
40 - 80% suhe tvari. Određuju najvažnija biološka svojstva bakterija i obično se sastoje od kombinacija 20 aminokiselina. Bakterije uključuju diaminopimelovu kiselinu (DAP), koje nema u stanicama ljudi i životinja. Bakterije sadrže više od 2000 različitih proteina koji se nalaze u strukturnim komponentama i uključeni su u metaboličke procese. Većina proteina ima enzimsko djelovanje.Proteini bakterijske stanice određuju antigenost i imunogenost, virulenciju i vrste bakterija. |
||
|
№ |
Element kompozicije |
Karakteristike i uloga u staničnoj fiziologiji. |
|
|
Nukleinske kiseline |
Oni obavljaju funkcije slične nukleinskim kiselinama eukariotskih stanica: molekula DNA u obliku kromosoma odgovorna je za nasljedstvo, ribonukleinske kiseline (informacijske ili matrične, transportne i ribosomske) uključene su u biosintezu proteina. |
||
|
Ugljikohidrati |
Predstavljeni su jednostavnim tvarima (mono- i disaharidi) i složenim spojevima. Polisaharidi se često nalaze u kapsulama. Neki unutarstanični polisaharidi (škrob, glikogen itd.) Su rezervne hranjive tvari. |
||
|
Lipidi |
Oni su dio citoplazmatske membrane i njezinih derivata, kao i stanične stjenke bakterija, na primjer, vanjske membrane, gdje osim biomolekularnog sloja lipida postoji i LPS. Lipidi mogu igrati ulogu rezervnih hranjivih tvari u citoplazmi. Bakterijske lipide predstavljaju fosfolipidi, masne kiseline i gliceridi. Mycobacterium tuberculosis sadrži najveću količinu lipida (do 40%). |
||
|
Minerali |
Nalazi se u pepelu nakon spaljivanja stanica. U velikim se količinama otkrivaju fosfor, kalij, natrij, sumpor, željezo, kalcij, magnezij, kao i elementi u tragovima (cink, bakar, kobalt, barij, mangan itd.) Uključeni su u regulaciju osmotskog tlaka, pH medija, redoks potencijal, aktiviraju enzime, dio su enzima, vitamina i strukturnih komponenti mikrobne stanice. |
||
Tablica 1.2. Dušične baze.
|
№ |
Dušične baze |
Karakteristično |
Bilješka |
|
Purin |
Adenin, Guanine |
Sastav nukleotida: deoksiriboza, dušične baze - adenin, gvanin, citozin, timin, ostatak H3PO4. Komplementarnost dušikovih baza A = T, G = C. Dvostruka spirala. Sposobni se udvostručiti |
|
|
Pirimidin |
Citozin, Timin ili Uracil (za RNK umjesto Timina) |
Tablica 1.2.1 Enzimi
|
№ |
Znak |
Karakteristično |
|
|
Definicija |
Specifični i učinkoviti proteinski katalizatori prisutni su u svim živim stanicama. |
||
|
Funkcije |
Enzimi smanjuju aktivacijsku energiju, osiguravajući pojavu takvih kemijskih reakcija koje bi se bez njih mogle odvijati samo pri visokim temperaturama, prekomjernom tlaku i pod drugim nefiziološkim uvjetima neprihvatljivim za živu stanicu. |
||
|
Enzimi povećavaju brzinu reakcije za oko 10 redova veličine, što smanjuje vrijeme poluraspada svake reakcije s 300 godina na jednu sekundu. |
|||
|
Enzimi "prepoznaju" supstrat po prostornom rasporedu njegove molekule i raspodjeli naboja u njoj. Određeni dio enzimske proteinske molekule - njezino katalitičko središte - odgovoran je za vezanje na supstrat. U tom slučaju nastaje intermedijarni kompleks enzim-supstrat, koji se zatim raspada stvaranjem produkta reakcije i slobodnog enzima. |
|||
|
Sorte |
Regulatorni (alosterični) enzimi percipiraju različite metaboličke signale i u skladu s njima mijenjaju svoju katalitičku aktivnost. |
Efektorski enzimi - enzimi koji kataliziraju određene reakcije (više detalja vidi u tablici 1.2.2.) |
|
|
Funkcionalna aktivnost |
Funkcionalna aktivnost enzima i brzina enzimskih reakcija ovise o uvjetima u kojima se mikroorganizam nalazi i, prije svega, o temperaturi medija i njegovom pH. Za mnoge patogene mikroorganizme optimalna temperatura je 37 ° C i pH 7,2-7,4. |
||
RAZRED ENZIMA:
-
mikroorganizmi sintetiziraju različite enzime koji pripadaju svih šest poznatih klasa.
Tablica 1.2.2. Klase efektorskih enzima
|
№ |
Klasa enzima |
Katalizuje: |
|
Oksidoreduktaza |
Prijenos elektrona |
|
|
Transferaze |
Prijenos različitih kemijskih skupina |
|
|
Hidrolaze |
Prijenos funkcionalnih skupina na molekulu vode |
|
|
Lijaze |
Pripajanje skupina na dvostruke veze i obrnute reakcije |
|
|
Izomeraza |
Prijenos skupina unutar molekule s stvaranjem izomernih oblika |
|
|
Ligase |
Stvaranje veza C-C, C-S, C-O, C-N zbog reakcija kondenzacije povezane s razgradnjom adenozin trifosfata (ATP) |
Tablica 1.2.3. Vrste enzima stvaranjem u bakterijskoj stanici
|
№ |
Vrsta |
Karakteristično |
Bilješke (uredi) |
|
Izvodljivo (prilagodljivo) enzima "Indukcija podloge" |
|
||
|
Represivni enzimi |
Sinteza ovih enzima je potisnuta kao posljedica prekomjerne akumulacije produkta reakcije koju katalizira ovaj enzim. |
Primjer potiskivanja enzima je sinteza triptofana koji nastaje iz antranilne kiseline uz sudjelovanje antranilat sintetaze. |
|
|
Konstitutivni enzimi |
Enzimi se sintetiziraju bez obzira na uvjete okoliša |
Enzimi glikolize |
|
|
Multienzimski kompleksi |
Unutarstanični enzimi spojeni strukturno i funkcionalno |
Enzimi respiratornog lanca lokalizirani na citoplazmatskoj membrani. |
Tablica 1.2.4. Specifični enzimi
|
№ |
Enzimi |
Identifikacija bakterija |
|
Superoksid dismutaza i katalaza |
Svi aerobi ili fakultativni anaerobi posjeduju superoksid dismutazu i katalazu - enzime koji štite stanicu od otrovnih produkata metabolizma kisika. Gotovo svi obligatni anaerobi ne sintetiziraju te enzime. Samo jedna skupina aerobnih bakterija - bakterije mliječne kiseline su katalaze negativne. |
|
|
Peroksidaza |
Bakterije mliječne kiseline akumuliraju peroksidazu - enzim koji katalizira oksidaciju organskih spojeva pod djelovanjem H2O2 (reducirano u vodu). |
|
|
Arginin dihidrolaza |
Dijagnostička značajka koja omogućuje razlikovanje saprofitnih vrsta Pseudomonas od fitopatogenih. |
|
|
Ureaza |
Među pet glavnih skupina obitelji Enterobacteriaceae, samo dvije - Escherichiae i Erwiniae - ne sintetiziraju ureazu. |
Tablica 1.2.5. Primjena bakterijskih enzima u industrijskoj mikrobiologiji.
|
№ |
Enzimi |
Primjena |
|
Amilaze, celulaze, proteaze, lipaze |
Za poboljšanje probave koriste se gotovi pripravci enzima koji olakšavaju hidrolizu škroba, celuloze, proteina i lipida. |
|
|
Invertaza kvasca |
U proizvodnji slatkiša za sprječavanje kristalizacije saharoze |
|
|
Pektinaza |
Koristi se za bistrenje voćnih sokova |
|
|
Kolagenaza klostridija i streptokinaza streptokoka |
Hidrolizira proteine, potiče zacjeljivanje rana i opeklina |
|
|
Litički enzimi bakterija |
Izlučuju se u okoliš, djeluju na stanične stijenke patogenih mikroorganizama i služe kao učinkovito sredstvo u borbi protiv potonjih, čak i ako imaju višestruku rezistenciju na antibiotike |
|
|
Ribonukleaze, deoksiribonukleaze, polimeraze, DNA ligaze i drugi enzimi koji specifično mijenjaju nukleinske kiseline |
Koristi se kao alat u bioorganskoj kemiji, genetskom inženjeringu i genskoj terapiji |
Tablica 1.2.6. Klasifikacija enzima prema lokalizaciji.
|
№ |
Klasa |
Lokalizacija |
Funkcije |
|
Endozimi |
|
Djeluju samo unutar stanice. Oni kataliziraju reakcije biosinteze i energetskog metabolizma. |
|
|
Egzocimi |
Puštaju se u okoliš. |
Stanica ih oslobađa u okoliš i katalizira reakcije hidrolize složenih organskih spojeva na jednostavnije koji su dostupni za asimilaciju od strane mikrobne stanice. Tu spadaju hidrolitički enzimi koji imaju iznimno važnu ulogu u prehrani mikroorganizama. |
Tablica 1.2.7.Enzimi patogenih mikroba (enzimi agresije)
|
№ |
Enzimi |
Funkcija |
Tvorba određenih enzima u laboratoriju |
|
Lecitovitellase = lecitinaza |
Uništava stanične membrane |
|
|
|
Hemolizin |
Uništava crvena krvna zrnca |
|
|
|
Kulture pozitivne na koagulazu |
Uzrokuje zgrušavanje krvne plazme |
|
|
|
Koagulaza negativne kulture |
Proizvodnja manitola |
|
|
|
№ |
Enzimi |
Funkcija |
Tvorba određenih enzima u laboratoriju |
|
Hijaluronidaza |
Hidrolizira hijaluronsku kiselinu - glavnu komponentu vezivnog tkiva |
|
|
|
Neuraminidaza |
Cijepa sialinsku (neuraminsku) kiselinu iz različitih glikoproteina, glikolipida, polisaharida, povećavajući propusnost različitih tkiva. |
Detekcija: reakcija za određivanje antitijela na neuraminidazu (RINA) i druga (imunodifuzija, imunoenzimske i radioimune metode). |
Tablica 1.2.8. Klasifikacija enzima prema biokemijskim svojstvima.
|
№ |
Enzimi |
Funkcija |
Otkrivanje |
|
Sugarolitički |
Razgradnja šećera |
Diferencijalno - dijagnostička okruženja kao što su Gissovo okruženje, Olkenitskovo okruženje, Endovo okruženje, Levinovo okruženje, Ploskirevo okruženje. |
|
|
Proteolitički |
Razgradnja proteina |
Mikrobi se inokuliraju injekcijom u kolonu želatine, a nakon 3-5 dana inkubacije na sobnoj temperaturi primjećuje se karakter ukapljivanja želatine. Proteolitička aktivnost određena je i stvaranjem produkata razgradnje proteina: indola, sumporovodika, amonijaka. Kako bi se odredili, mikroorganizmi se inokuliraju u mesno-peptonsku juhu. |
|
|
Enzimi krajnjih proizvoda |
|
Koriste se za razlikovanje nekih vrsta bakterija od drugih na temelju njihove enzimske aktivnostiokruženja za diferencijalnu dijagnostiku |
Shema 1.2.8. Sastav enzima.
ENZIMSKI SASTAV BILO KOG MIKROORGANIZMA:
Definirano svojim genomom
Je li stabilan znak
Često se koristi za njihovu identifikaciju
Određivanje saharolitičkih, proteolitičkih i drugih svojstava.
Tablica 1.3. Pigmenti
|
№ |
Pigmenti |
Sinteza mikroorganizama |
|
Karotenoidni pigmenti topljivi u masti u crvenoj, narančastoj ili žutoj boji |
Formiraju sarcine, mycobacterium tuberculosis, neke aktinomicete. Ovi pigmenti štite ih od UV zraka. |
|
|
Crni ili smeđi pigmenti - melanini |
Sintetizirani obligatnim anaerobima Bacteroides niger i drugima, netopivi u vodi, pa čak i jakim kiselinama |
|
|
Svijetlocrveni pigment pirola - prodigiosin |
Formirana nekim seracijama |
|
|
Fenozin pigment topiv u vodi - piocijanin. |
Proizvode ga bakterije Pseudomonas aeruginosa (Pseudomonas aeruginosa). U tom slučaju medij za kulturu s neutralnim ili alkalnim pH postaje plavo-zelen. |
Tablica 1.4. Svjetlosni mikroorganizmi koji stvaraju aromu
|
№ |
Fenomen |
Stanje i karakteristike |
|
Sjaj (luminescencija) |
Bakterije uzrokuju luminiscenciju tih supstrata, na primjer, ljuske ribe, veće gljive, trula stabla, prehrambeni proizvodi, na čijoj se površini razmnožavaju.Većina luminiscentnih bakterija su halofilne vrste koje se mogu razmnožavati pri povišenim koncentracijama soli. Žive u morima i oceanima, a rijetko u slatkovodnim tijelima. Sve luminiscentne bakterije su aerobne. Mehanizam luminiscencije povezan je s oslobađanjem energije tijekom biološke oksidacije supstrata. |
|
|
Formiranje arome |
Neki mikroorganizmi proizvode hlapljive arome, poput etil acetata i amil octenog estera, koji daju aromu vinu, pivu, mliječnoj kiselini i drugim prehrambenim proizvodima, pa se stoga koriste u njihovoj proizvodnji. |
Tablica 2.1.1 Metabolizam
|
№ |
Koncept |
Definicija |
|
|
Metabolizam |
Biokemijski procesi u stanici ujedinjeni su jednom riječju - metabolizam (grč. Metabole - transformacija). Ovaj je izraz ekvivalentan konceptu "metabolizma i energije". Postoje dva aspekta metabolizma: anabolizam i katabolizam. |
||
|
|
||
|
Amfibolizam |
Intermedijarni metabolizam koji pretvara fragmente hranjivih tvari niske molekularne mase u brojne organske kiseline i estere fosfora naziva se |
||
Shema 2.1.1. Metabolizam
METABOLIZAM -
skup dva suprotna, ali međusobno povezana procesa: katabolizam i anabolizam
s
Anabolizam= asimilacija = plastični metabolizam = konstruktivni metabolizam
Katabolizam= disimilacija = energetski metabolizam = raspadanje = opskrba stanice energijom
Sinteza (sastavnica stanica)
Enzimske kataboličke reakcije rezultiraju oslobađanje energije, koji se nakupio u molekulama ATP -a.
Biosinteza monomera:
aminokiseline nukleotidi monosaharidi masnih kiselina
Biosinteza polimera:
proteini nukleinske kiseline, lipidni polisaharidi
Kao rezultat enzimske anaboličke reakcije, energija oslobođena u procesu katabolizma troši se na sintezu makromolekula organskih spojeva iz kojih se zatim sastavljaju biopolimeri - komponente mikrobne stanice.
Energija se troši na sintezu staničnih komponenti
Tablica 2.1.3. Metabolizam i transformacija stanične energije.
|
№ |
Metabolizam |
Karakteristično |
Bilješke (uredi) |
|
Funkcija |
Metabolizam osigurava dinamičku ravnotežu svojstvenu živom organizmu kao sustavu, u kojem su sinteza i uništavanje, reprodukcija i smrt međusobno uravnotežene. |
Metabolizam je glavni znak života |
|
|
Razmjena plastike Sinteza proteina, masti, ugljikohidrata. |
Ovo je skup reakcija biološke sinteze. |
Od tvari koje izvana ulaze u stanicu nastaju molekule, slične spojevima stanice, odnosno dolazi do asimilacije. |
|
|
Razmjena energije propadanje |
Proces je suprotan sintezi. Ovo je zbirka reakcija cijepanja. |
Prilikom razdvajanja visokomolekularnih spojeva oslobađa se energija koja je neophodna za reakciju biosinteze, odnosno dolazi do disimilacije.
|
Tablica 2.1.2. Razlika u metabolizmu radi identifikacije.
|
№ |
Opcije |
|
Sposobnost korištenja različitih tvari kao izvora ugljika. |
|
|
Sposobnost stvaranja specifičnih krajnjih proizvoda kao rezultat razgradnje podloga. |
|
|
Sposobnost miješanja pH medija za uzgoj na kiselu ili alkalnu stranu.Metabolizam većine bakterija provodi se biokemijskim reakcijama razgradnje organskih (rjeđe anorganskih) tvari i sintezom komponenata bakterijskih stanica iz jednostavnih spojeva koji sadrže ugljik. |
Tablica 2.2 Anabolizam (konstruktivni metabolizam)
|
№ |
Anabolička reakcijska skupina |
Sintetizirano: |
|
Biosinteza monomera |
Aminokiseline, nukleotidi, monosaharidi, masne kiseline |
|
|
Biosinteza polimera |
Proteini, nukleinske kiseline, polisaharidi i lipidi |
Shema 2.2.2. Biosinteza aminokiselina u prokariota.
Autor - L.B. Borisov, str. 52 "Medicinska mikrobiologija"

Shema 2.2.1. Biosinteza ugljikohidrata u mikroorganizmima.
Autor - L.B. Borisov, p. 51 "Medicinska mikrobiologija"
Slika 2.2.3. Biosinteza lipida
Tablica 2.2.4. Faze metabolizma energije - katabolizam.
|
№ |
Faze |
Karakteristično |
Bilješka |
|
Pripremna |
Molekule disaharida i polisaharida, proteini se razgrađuju na male molekule - glukozu, glicerin i masne kiseline, aminokiseline. Velike molekule nukleinske kiseline po nukleotidu. |
U ovoj fazi oslobađa se mala količina energije koja se rasipa u obliku topline. |
|
|
Anoksično ili nepotpuno ili anaerobno ili fermentirano ili disimilirano. |
Tvari nastale u ovoj fazi uz sudjelovanje enzima dodatno se razgrađuju. Na primjer: glukoza se razlaže na dvije molekule mliječne kiseline i dvije molekule ATP -a. |
ATP i H3PO4 sudjeluju u reakcijama cijepanja glukoze. Tijekom razgradnje glukoze bez kisika, 40% energije pohranjeno je u molekuli ATP-a u obliku kemijske veze, ostatak se rasipa u obliku topline. U svim slučajevima raspada jedne molekule glukoze nastaju dvije molekule ATP. |
|
|
Faza aerobnog disanja ili razgradnje kisika. |
Kad je stanici dostupan kisik, tvari nastale tijekom prethodne faze oksidiraju se (razgrađuju) do konačnih proizvodaCO i HO.. |
Ukupna jednadžba aerobnog disanja:
|
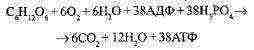
Shema 2.2.4. Vrenje.
Fermentacijski metabolizam -karakterizirana stvaranjem ATP -a fosforilacijom podloga.
-
Prvo (oksidacija) = cijepanje
-
Drugo (oporavak)
Uključuje pretvorbu glukoze u piruvičnu kiselinu.
Uključuje oporabu vodika za oporabu piruvične kiseline.
Putevi za stvaranje piruvične kiseline iz ugljikohidrata
Shema 2.2.5. Piruvična kiselina.
Glikolitički put (Embden-Meyerhof-Parnassus put)
Staza Entner-Dudorov
Pentozni fosfatni put
Tablica 2.2.5. Vrenje.
|
№ |
Vrsta fermentacije |
Predstavnici |
Finalni proizvod |
Bilješke (uredi) |
|
Mliječna kiselina |
|
Od piruvata tvori mliječnu kiselinu |
U nekim slučajevima (homofermentna fermentacija) nastaje samo mliječna kiselina, u drugim također nusproizvodi. |
|
|
Mravlja kiselina |
|
Mravlja kiselina jedan je od krajnjih proizvoda. (zajedno s njom - strana) |
Neke vrste Enterobacteriaceae razgrađuju mravlju kiselinu na H2 i CO2 / |
|
|
Maslačna kiselina |
|
Maslačna kiselina i nusproizvodi |
Neke vrste klostridija, zajedno s maslačnom i drugim kiselinama, tvore butanol, aceton itd. (Tada se naziva aceton-butilna fermentacija). |
|
|
Propionska kiselina |
|
Od piruvata tvori propionsku kiselinu |
Mnoge bakterije fermentiraju ugljikohidrate zajedno s drugom hranom i tvore etilni alkohol. Međutim, to nije glavni proizvod. |
Tablica 2.3.1. Sustav sinteze proteina, ionska izmjena.
|
№ |
Ime proizvoda |
Karakteristično |
|
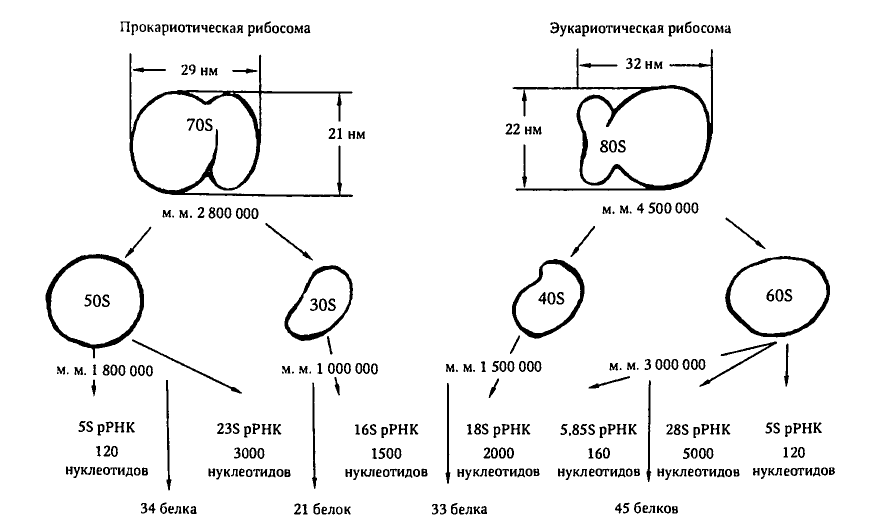
Ribosomske podjedinice 30S i 50S |
U slučaju bakterijskih 70S ribosoma, 50S podjedinica sadrži 23S rRNA (~ 3000 nukleotida duljine), a 30S podjedinica sadrži 16S rRNA (~ 1500 nukleotida duljine); velika ribosomska podjedinica, osim "duge" rRNA, sadrži i jednu ili dvije "kratke" rRNA (5S rRNA bakterijskih ribosomskih podjedinica 50S ili 5S i 5,8S rRNA velikih ribosomskih podjedinica eukariota). (Za više detalja pogledajte sliku 2.3.1.) |
|
|
Messenger RNA (mRNA) |
RNA koja sadrži informacije o primarnoj strukturi (aminokiselinski slijed) proteina |
|
|
Kompletan skup od dvadeset aminoacil-tRNA, za čije su stvaranje potrebne odgovarajuće aminokiseline, aminoacil-tRNA sintetaze, tRNA i ATP |
To je aminokiselina, napunjena energijom i vezana za tRNA, spremna za transport do ribosoma i ugrađena u polipeptid koji se na njemu sintetizira. |
|
|
Transportna RNA (tRNA) |
Ribonukleinska kiselina, čija je funkcija transport aminokiselina do mjesta sinteze proteina. |
|
|
Faktori pokretanja proteina |
(u prokariotima-IF-1, IF-2, IF-3) Dobili su ime jer su uključeni u organizaciju aktivnog kompleksa (708-kompleks) od 30S i 50S podjedinica, mRNA i inicijatorske aminoacil-tRNA (u prokarioti - formilmetionil -tRNA), koji "pokreće" (pokreće) rad ribosoma - prijevod mRNA. |
|
|
Čimbenici produljenja proteina |
(u prokariota-EF-Tu, EF-Ts, EF-G) Sudjelujte u produženju (produženju) sintetiziranog polipeptidnog lanca (peptidil). Faktori oslobađanja proteina (RF) omogućuju kodonu specifično odvajanje polipeptida od ribosoma i završetak sinteze proteina. |
|
|
№ |
Ime proizvoda |
Karakteristično |
|
Faktori prekidanja proteina |
(u prokariota-RF-1, RF-2, RF-3) |
|
|
Neki drugi proteinski čimbenici (asocijacije, disocijacija podjedinica, otpuštanje itd.). |
Faktori prevođenja proteina potrebni za funkcioniranje sustava |
|
|
Guanozin trifosfat (GTP) |
Za emitiranje je potrebno sudjelovanje GTF -a. Potreba sustava za sintezu proteina za GTP vrlo je specifična: ne može se zamijeniti niti jednim od drugih trifosfata. Stanica troši više energije na biosintezu proteina nego na sintezu bilo kojeg drugog biopolimera. Za stvaranje svake nove peptidne veze potrebno je cijepanje četiri visokoenergetske veze (ATP i GTP): dvije kako bi se molekula tRNA opteretila aminokiselinom, te još dvije tijekom produljenja-jedna tijekom vezanja aa-tRNA, a druga tijekom translokacije. |
|
|
Anorganski kationi pri određenoj koncentraciji. |
Za održavanje pH sustava u fiziološkim granicama. Amonijeve ione neke bakterije koriste za sintezu aminokiselina, kalijeve ione za vezanje tRNA na ribosome. Ioni željeza i magnezija igraju ulogu kofaktora u brojnim enzimskim procesima |
Slika 2.3.1. Shematski prikaz struktura prokariotskih i eukariotskih ribosoma.
Autor - Korotyaev, str. 68 "Medicinska mikrobiologija"
Tablica 2.3.2. Značajke ionske izmjene u bakterijama.
|
№ |
Osobitost |
Karakterizira: |
|
|
Visok osmotski tlak |
Zbog značajne unutarstanične koncentracije kalijevih iona u bakterijama održava se visoki osmotski tlak. |
||
|
Unos željeza |
Za brojne patogene i oportunističke bakterije (Escherichia, Shigella itd.), Potrošnja željeza u tijelu domaćina otežana je zbog njegove netopivosti pri neutralnim i blago alkalnim pH vrijednostima. |
Siderofori -posebne tvari koje vezivanjem željeza čine topljivim i transportirajućim. |
|
|
Asimilacija |
Bakterije aktivno asimiliraju anione SO2 / i PO34 + iz okoliša radi sinteze spojeva koji sadrže te elemente (aminokiseline koje sadrže sumpor, fosfolipide itd.). |
||
|
Jona |
Za rast i razmnožavanje bakterija potrebni su mineralni spojevi - ioni NH4 +, K +, Mg2 +itd. (Za više detalja vidjeti tablicu 2.3.1.) |
||
Tablica 2.3.3. Ionska izmjena
|
№ |
Naziv mineralnih spojeva |
Funkcija |
|
NH4 + (amonijevi ioni) |
Koriste ga neke bakterije za sintezu aminokiselina |
|
|
K + (kalijevi ioni) |
|
|
|
Fe2 + (ioni željeza) |
|
|
|
Mg2 + (magnezijevi ioni) |
||
|
SO42- (sulfatni anion) |
Potrebno za sintezu spojeva koji sadrže te elemente (aminokiseline koje sadrže sumpor, fosfolipide itd.) |
|
|
PO43- (fosfatni anion) |
Shema 2.4.1. Energetski metabolizam.
Za sintezu bakterijama je potrebno ...
-
Hranjive tvari
-
Energija
Tablica 2.4.1. Energetski metabolizam (biološka oksidacija).
|
№ |
Postupak |
Potrebno: |
|
Sinteza strukturnih komponenti mikrobnih stanica i održavanje vitalnih procesa |
Odgovarajuća količina energije. Ta se potreba zadovoljava biološkom oksidacijom, zbog čega se sintetiziraju molekule ATP. |
|
|
Energija (ATP) |
Željezne bakterije primaju energiju oslobođenu tijekom izravne oksidacije željeza (Fe2 + u Fe3 +), koja se koristi za fiksiranje CO2, bakterije koje metaboliziraju sumpor, opskrbljuju se energijom zbog oksidacije spojeva koji sadrže sumpor. Međutim, velika većina prokariota dobiva energiju dehidrogenacijom. Energija se također prima u procesu disanja (za detaljnu tablicu pogledajte odgovarajući odjeljak). |
Shema 2.4. Biološka oksidacija u prokariota.
Razlaganje polimera u monomere
Faza I.
Protein
Masti
Ugljikohidrati
glicerin i masne kiseline
aminokiseline
monosaharidi
Razlaganje u anoksičnim uvjetima
Faza II
Formiranje međuproizvoda
Oksidacija u uvjetima kisika do konačnih proizvoda
Faza III
CO2
H20
Tablica 2.4.2. Energetski metabolizam.
|
№ |
Koncept |
Karakteristično |
|
Bit energetskog metabolizma |
Pružanje energije stanicama potrebne za očitovanje života. |
|
|
ATF |
Molekula ATP -a sintetizirana je kao rezultat prijenosa elektrona s primarnog davatelja na konačni akceptor. |
|
|
Dah |
|
|
|
Mobilizacija energije |
Energija se mobilizira u oksidacijskim i redukcijskim reakcijama. |
|
|
Reakcija oksidacije |
Sposobnost tvari da donira elektrone (oksidira) |
|
|
Reakcija oporavka |
Sposobnost tvari da veže elektrone. |
|
|
Redoks potencijal |
Sposobnost tvari da donira (oksidira) ili prima (oporavi) elektrone. (kvantitativni izraz) |
Shema 2.5. Sinteza.
SINTEZA
bjelančevine
mast
ugljikohidrata
Tablica 2.5.1. Sinteza
|
№ |
Ime |
Karakteristično |
|
Citoplazma |
Sinteza početnih produkata odvija se u citoplazmi. |
|
|
Citoplazmatska membrana |
Početni proizvodi iz citoplazme prenose se na vanjsku površinu citoplazmatske membrane. |
|
|
Morfogeneza |
Na CPM -u započinje morfogeneza, odnosno stvaranje staničnih struktura (kapsula, staničnih stjenki itd.) Uz sudjelovanje enzima. |
Tablica 2.5.1. Sinteza
|
№ |
Biosinteza |
Ili što |
Bilješke (uredi) |
|
Ja |
Biosinteza ugljikohidrata |
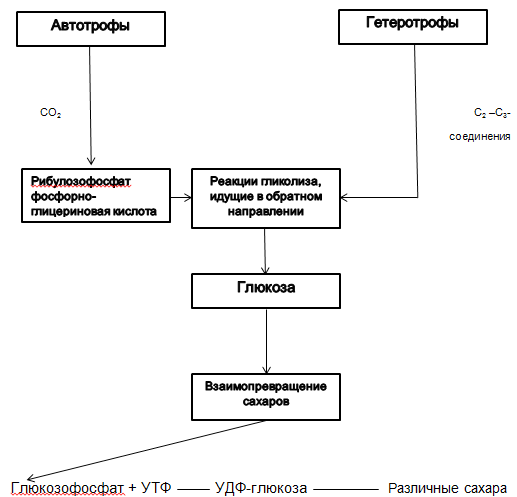
Autotrofi sintetiziraju glukozu iz CO2. Heterotrofi sintetiziraju glukozu iz spojeva koji sadrže ugljik. |
Calvinov ciklus (vidi dijagram 2.2.1.) |
|
II |
Biosinteza aminokiselina |
Većina prokariota može sintetizirati sve aminokiseline iz:
|
Izvor energije je ATP. Piruvat nastaje u glikolitičkom ciklusu. Auksotrofni mikroorganizmi - konzumiraju se gotovi u tijelu domaćina. |
|
III |
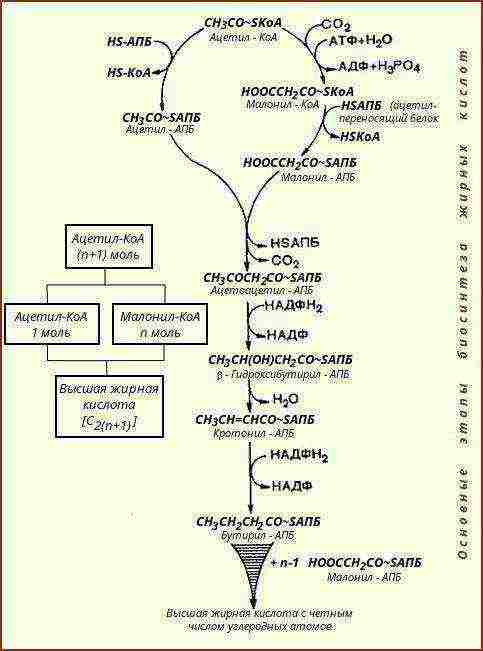
Biosinteza lipida |
Lipidi se sintetiziraju iz jednostavnijih spojeva - metaboličkih produkata proteina i ugljikohidrata |
Proteini za prijenos acetil igraju važnu ulogu. Auksotrofni mikroorganizmi - konzumirajte gotove u tijelu domaćina ili iz hranjivih medija. |
Tablica 2.5.2. Glavne faze biosinteze proteina.
|
№ |
Faze |
Karakteristično |
Bilješke (uredi) |
|
Transkripcija |
Proces sinteze RNA na genima. Ovo je proces prepisivanja informacija iz gena DNA - gena u mRNA - gena. |
Provodi se pomoću DNK -ovisne RNA -polimeraze. Prijenos informacija o strukturi proteina na ribosome događa se uz pomoć mRNA. |
|
|
Emitiranje (prijenos) |
Proces biosinteze vlastitih proteina. Postupak dekodiranja genetskog koda u mRNA i njegova implementacija u obliku polipeptidnog lanca. |
Budući da svaki kodon sadrži tri nukleotida, isti se genetski tekst može čitati na tri različita načina (počevši od prvog, drugog i trećeg nukleotida), to jest u tri različita okvira za čitanje. |
-
Napomena u tablici: Primarna struktura svakog proteina je niz aminokiselina u njemu.
Shema 2.5.2. Lanci prijenosa elektrona od primarnog donatora vodika (elektroni) do njegovog konačnog akceptora O2.
Organska tvar
(primarni donator elektrona)
NAD (- 0,32)
Flavoprotein (- 0,20)
Kinon (- 0, 07)
Citokrom (+0,01)
Citokrom C (+0,22)
Citokrom A (+0,34)
O2 (+0,81)
konačni akceptor
Tablica 3.1. Razvrstavanje organizama prema vrstama hrane.
|
№ |
Organogeni element |
Vrste hrane |
Karakteristično |
|
Ugljik (C) |
|
Oni sami sintetiziraju sve komponente stanice koje sadrže ugljik iz CO2. |
|
|
Ne mogu zadovoljiti svoje potrebe CO2, koriste gotove organske spojeve. |
||
|
Izvor hrane su mrtve organske podloge. |
||
|
Izvor hrane su živa tkiva životinja i biljaka. |
||
|
Dušik (N) |
|
Zadovoljiti njihove potrebe atmosferskim i mineralnim dušikom |
|
|
Potrebni su gotovi organski dušikovi spojevi. |
||
|
Vodik (H) |
Glavni izvor je H2O |
||
|
Kisik (O) |
|||
Tablica 3.1.2. Transformacija energije
|
№ |
Klasifikacija |
Ime |
Potreban: |
|
Po izvoru energije |
|
sunčevu svjetlost |
|
|
Redoks reakcije |
||
|
Donatorom elektrona |
|
Anorganski spojevi (H2, H2S, NH3, Fe itd.) |
|
|
Organski spojevi |
Tablica 3.1.3. Metode hranjenja ugljikom
|
№ |
Izvor energije |
Donator elektrona |
Način hranjenja ugljikom |
|
Energija sunčeve svjetlosti |
Anorganski spojevi |
Fotolitoheterotrofi |
|
|
Organski spojevi |
Fotoorganoheterotrofi |
||
|
Redoks reakcije |
Anorganski spojevi |
Kemolithoheterotrofi |
|
|
Organski spojevi |
Kemoorganoheterotrofi |
Tablica 3.2. Mehanizmi napajanja:
|
№ |
Mehanizam |
Uvjeti |
Gradijent koncentracije |
Troškovi energije |
Specifičnost podloge |
|
Pasivna difuzija |
Koncentracija hranjivih tvari u mediju prelazi koncentraciju u stanici. |
Gradijentom koncentracije |
– |
– |
|
|
Olakšana difuzija |
Uključeni su proteini permeaze. |
Gradijentom koncentracije |
– |
+ |
|
|
Aktivni transport |
Uključeni su proteini permeaze. |
Protiv gradijenta koncentracije |
+ |
+ |
|
|
3A |
Translokacija kemijskih skupina |
Tijekom procesa prijenosa dolazi do kemijske modifikacije hranjivih tvari. |
Protiv gradijenta koncentracije |
+ |
+ |
Tablica 3.3. Transport hranjivih tvari iz bakterijske stanice.
|
№ |
Ime |
Karakteristično |
|
Reakcija fosfotransferaze |
Javlja se pri fosforilaciji prenesene molekule. |
|
|
Translacijsko lučenje |
U tom slučaju sintetizirane molekule moraju imati posebnu vodeću aminokiselinsku sekvencu kako bi se spojile na membranu i formirale kanal kroz koji proteinske molekule mogu pobjeći u okoliš. Tako se iz stanica odgovarajuće bakterije oslobađaju toksini tetanusa, difterije i drugih molekula. |
|
|
Pupanje membrane |
Molekule nastale u stanici okružene su membranskom vezikulom, koja se odvaja u okoliš. |
Tablica 4. Rast.
|
№ |
Koncept |
Definicija pojma. |
|
Visina |
Nepovratno povećanje količine žive tvari, najčešće zbog diobe stanica.Ako se u višestaničnim organizmima obično uočava povećanje tjelesne veličine, tada se u višestaničnim organizmima povećava broj stanica. No čak i kod bakterija treba razlikovati povećanje broja stanica i povećanje stanične mase. |
|
|
Čimbenici koji utječu na rast bakterija in vitro. |
Mycobacterium leprae nije sposoban in vitro Rast klamidije (uključujući parazite)
|
|
|
Procjena rasta bakterija |
Kvantificiranje rasta obično se provodi u tekućim medijima gdje rastuće bakterije tvore homogenu suspenziju. Povećanje broja stanica utvrđuje se određivanjem koncentracije bakterija u 1 ml, ili se povećanje stanične mase određuje u jedinicama mase po jedinici volumena. |
Čimbenici rasta
Lipidi
Aminokiseline
Vitamini
Dušične baze
Tablica 4.1. Čimbenici rasta
|
№ |
Čimbenici rasta |
Karakteristično |
Funkcija |
|
|
Aminokiseline |
|
Mnogi mikroorganizmi, osobito bakterije, trebaju jednu ili više aminokiselina (jednu ili više), budući da ih ne mogu sami sintetizirati. Mikroorganizmi ove vrste nazivaju se auksotrofni za one aminokiseline ili druge spojeve koje ne mogu sintetizirati. |
||
|
Purinske baze i njihovi derivati |
Nukleotidi:
|
Oni su čimbenici rasta bakterija. Neke vrste mikoplazmi trebaju nukleotide. Potrebno za izgradnju nukleinskih kiselina. |
||
|
Pirimidinske baze i njihovi derivati |
Nukleotidi
|
|||
|
№ |
Čimbenici rasta |
Karakteristično |
Funkcija |
|
|
Lipidi |
|
Dio membranskih lipida |
||
|
||||
|
Jesu li komponente fosfolipida |
|||
|
U mikoplazmama su dio citoplazmatske membrane |
|||
|
||||
|
Vitamini (uglavnom skupina B) |
|
Staphylococcus aureus, pneumokok, Brucella |
||
|
Sve vrste bakterija u obliku štapića |
|||
|
Bifidobakterije i propionska kiselina |
|||
|
Neke vrste streptokoka, bacili tetanusa |
|||
|
Kvasci i bakterije koje učvršćuju dušik Rhizobium |
|||
|
Heme - komponente citokroma |
Hemofilne bakterije, mikobakterija tuberkuloza |
|||
Tablica 5. Disanje.
|
№ |
Ime |
Karakteristično |
|
Dah |
Biološka oksidacija (enzimske reakcije) |
|
|
Baza |
Disanje se temelji na redoks reakcijama koje dovode do stvaranja ATP -a, univerzalnog akumulatora kemijske energije. |
|
|
Procesi |
Prilikom disanja događaju se sljedeći procesi:
|
|
|
Aerobno disanje |
Konačni akceptor vodika ili elektrona je molekularni kisik. |
|
|
Anaerobno disanje |
Akceptor vodika ili elektrona je anorganski spoj-NO3-, SO42-, SO32-. |
|
|
Vrenje |
Organski spojevi su akceptori vodika ili elektrona. |
Tablica 5.1. Klasifikacija disanja.
|
№ |
Bakterije |
Karakteristično |
Bilješke (uredi) |
|
Strogi anaerobi |
|
|
|
|
Strogi aerobi |
|
Strogi aerobi uključuju, na primjer, predstavnike roda Pseudomonas |
|
|
№ |
Bakterije |
Karakteristično |
Bilješke (uredi) |
|
Fakultativni anaerobi |
|
Fakultativni anaerobi uključuju enterobakterije i mnoge kvasce koji mogu prijeći s disanja u prisutnosti O2 na fermentaciju u odsutnosti O2. |
|
|
Mikroaerofili |
Mikroorganizam koji zahtijeva, za razliku od strogih anaeroba, prisutnost kisika u atmosferi ili hranjivom mediju za svoj rast, ali u smanjenim koncentracijama u usporedbi sa sadržajem kisika u običnom zraku ili u normalnim tkivima tijela domaćina (za razliku od aeroba , za čiji rast je normalan sadržaj kisika u atmosferi ili hranjivom mediju). Mnogi mikroaerofili su također kapnofili, odnosno zahtijevaju povećanu koncentraciju ugljičnog dioksida. |
U laboratoriju se takvi organizmi lako uzgajaju u "staklenci za svijeće". "Staklenka za svijeće" je spremnik u koji se unosi goruća svijeća prije nego što se zatvori hermetičkim poklopcem. Plamen svijeće gorjet će sve dok se ne ugasi zbog nedostatka kisika, zbog čega se u limenci stvara atmosfera zasićena ugljikovim dioksidom sa smanjenim sadržajem kisika. |
Tablica 6. Značajke razmnožavanja.
|
№ |
Ime |
Karakteristično |
|
Reprodukcija |
Izraz "razmnožavanje" odnosi se na povećanje broja stanica u populaciji. Većina se prokariota razmnožava poprečnom diobom, neki pupanjem. Gljive se razmnožavaju sporulacijom. |
|
|
Kamo ide |
Kad se mikrobna stanica umnoži, najvažniji procesi događaju se u jezgri (nukleoid), koja sadrži sve genetske informacije u dvolančanoj molekuli DNA. |
Shema 6. Ovisnost trajanja proizvodnje o različitim čimbenicima.
Trajanje generacije
Vrsta bakterija
Dob
Populacija
Temperatura
Sastav hranjivog medija
Tablica 6.1. Faze razmnožavanja bakterija.
|
№ |
Faza |
Karakteristično |
|
Ja |
Početna stacionarna faza |
Traje 1-2 sata. Tijekom ove faze broj bakterijskih stanica se ne povećava. |
|
II |
Lag faza (faza odgode reprodukcije) |
Karakterizira ga početak intenzivnog rasta stanica, ali stopa stanične diobe ostaje niska. |
|
III |
Dnevnička faza (logaritamska) |
Razlikuje se maksimalnom brzinom reprodukcije stanica i povećanjem broja bakterijske populacije eksponencijalno |
|
IV |
Negativna faza ubrzanja |
Karakterizira ga niža aktivnost bakterijskih stanica i produljenje razdoblja stvaranja. To se događa kao posljedica iscrpljivanja hranjivog medija, nakupljanja metaboličkih proizvoda u njemu i nedostatka kisika. |
|
V. |
Stacionarna faza |
Karakterizira ga ravnoteža između broja mrtvih, novonastalih i uspavanih stanica. |
|
VI |
Faza propasti |
Javlja se konstantnom brzinom i zamjenjuje se UP-USH fazama smanjenja stope stanične smrti. |
Shema 7. Zahtjevi za kulturne medije.
Zahtjevi
Viskoznost
Vlažnost
Sterilnost
Nutritivna vrijednost
Transparentnost
Izotoničnost
pH okoliša
Tablica 7. Razmnožavanje bakterija na hranjivim podlogama.
|
№ |
Hranjivi medij |
Karakteristično |
|
|
Gusti hranjivi mediji |
Na gustim hranjivim medijima bakterije stvaraju kolonije - nakupine stanica. |
||
|
S - vrsta (glatko - glatko i sjajno) Okrugla, s ravnim rubom, glatka, ispupčena. |
R - vrsta (grubo - grubo, neujednačeno) Nepravilnog oblika sa nazubljenim rubovima, hrapavim, udubljenim. |
||
|
Tekući medij za kulturu |
|
||
Tablica 7.1. Klasifikacija kulturnih medija.
|
№ |
Klasifikacija |
Pogledi |
Primjeri |
|
Po sastavu |
Jednostavan |
|
|
|
Kompleks |
|
||
|
Po dogovoru |
Glavni |
|
|
|
Izborni |
|
||
|
Diferencijalno - dijagnostički |
|
||
|
Posebna |
|
||
|
Dosljednošću |
Gusta |
|
|
|
Tekući |
|
||
|
Polutekući |
|
||
|
Po podrijetlu |
Prirodno |
|
|
|
Polusintetičko |
|
||
|
Sintetičko |
|
Tablica 7.2. Načela izolacije čiste stanične kulture.
|
Mehanički princip |
Biološki princip |
|
METODE 1. Razlomljena razrjeđenja L. Pasteur 2. Razrjeđivanje ploča R. Koch 3. Površinski usjevi Drigalsky 4. Površinski potezi |
METODE Smatrati: a - vrsta disanja (Fortnerova metoda); b - pokretljivost (Šukevićeva metoda); c - otpornost na kiselinu; d - sporulacija; d - temperaturni optimum; e - selektivna osjetljivost laboratorijskih životinja na bakterije |
Tablica 7.2.1. Faze izolacije čiste stanične kulture.
|
№ |
Pozornica |
Karakteristično |
|
Faza 1 istraživanja |
Odnesite patološki materijal. Proučava se - izgled, konzistencija, boja, miris i drugi znakovi, priprema se razmaz, boji i pregledava pod mikroskopom. |
|
|
Faza 2 istraživanja |
Na površini guste hranjive podloge mikroorganizmi stvaraju kontinuirani, gusti rast ili izolirane kolonije.Kolonija - To su nakupine bakterija vidljive golim okom na površini ili u debljini hranjivog medija. U pravilu se svaka kolonija tvori od potomaka jedne mikrobne stanice (klonova), pa je njihov sastav prilično homogen. Značajke rasta bakterija na hranjivim podlogama očitovanje su njihovih kulturnih svojstava. |
|
|
Faza 3 istraživanja |
Proučava se priroda rasta čiste kulture mikroorganizama i provodi njezina identifikacija. |
Tablica 7.3. Identifikacija bakterija.
|
№ |
Ime |
Karakteristično |
|
Biokemijska identifikacija |
Određivanje vrste patogena prema njegovim biokemijskim svojstvima |
|
|
Serološka identifikacija |
Kako bi se ustanovile vrste bakterija, često se proučava njihova antigenska struktura, odnosno identificiraju se po antigenim svojstvima. |
|
|
Identifikacija po biološkim svojstvima |
Ponekad se bakterije identificiraju zaražavanjem laboratorijskih životinja čistom kulturom i promatranjem promjena koje patogeni uzrokuju u tijelu. |
|
|
Kulturna identifikacija |
Određivanje vrste patogena prema njihovim kulturnim karakteristikama |
|
|
Morfološka identifikacija |
Određivanje vrste bakterija prema njihovim morfološkim karakteristikama |
Kontrolni testovi ocjenjivanja
-
Koji od procesa nije povezan s fiziologijom bakterija?
-
Rast
-
Reprodukcija
-
Mutacija
-
Prehrana
-
Koje tvari čine 40 - 80% suhe mase bakterijske stanice?
-
Ugljikohidrati
-
Protein
-
Masti
-
Nukleinske kiseline
-
Koje klase enzima sintetiziraju mikroorganizmi?
-
Oksi reduktaza
-
Svi razredi
-
Transferaze
-
Ligase
-
Enzimi čija se koncentracija u stanici naglo povećava kao odgovor na pojavu supstrata induktora u mediju?
-
Izvodljivo
-
Ustavni
-
Represivno
-
Multienzimski kompleksi
-
Enzim patogenosti koji luči Staphylococcus aureus?
-
Neuraminidaza
-
Hijaluronidaza
-
Lecitinaza
-
Fibrinolizin
-
Obavljaju li proteolitički enzimi neku funkciju?
-
Razgradnja proteina
-
Razbijanje masti
-
Razgradnja ugljikohidrata
-
Stvaranje lužine
-
Fermentacija Enterobacteriaceae?
-
Mliječna kiselina
-
Mravlja kiselina
-
Propionska kiselina
-
Maslačna kiselina
-
Koji se mineralni spojevi koriste za vezanje t-RNA na ribosome?
-
NH4
-
K +
-
Fe2 +
-
Mg2 +
-
Biološka oksidacija je ...?
-
Prehrana
-
Reprodukcija
-
Dah
-
Stanična smrt
-
Koje tvari same sintetiziraju sve komponente stanice koje sadrže ugljik iz CO2.
-
Prototrofi
-
Heterotrofi
-
Autotrofi
-
Saprofiti
-
Mediji kulture razlikuju se:
-
Po sastavu
-
Dosljednošću
-
Po dogovoru
-
Sve od navedenog
-
Reproduktivna faza, koju karakterizira ravnoteža između broja mrtvih, novonastalih i uspavanih stanica?
-
Lag faza
-
Faza dnevnika
-
Negativna faza ubrzanja
-
Stacionarna faza
-
Ovisno o trajanju proizvodnje?
-
Vrsta
-
Dob
-
Populacije
-
Sve od navedenog
-
Kako bi se ustanovile vrste bakterija, često se proučava njihova antigenska struktura, odnosno provodi se identifikacija, koja?
-
Biološki
-
Morfološki
-
Serološki
-
Biokemijski
-
Način površinske sjetve Drygalskog naziva se ...?
-
Mehanički principi izolacije čiste kulture
-
Biološki principi izolacije čiste kulture
Bibliografija
1. Borisov LB Medicinska mikrobiologija, virologija, imunologija: udžbenik za med. sveučilišta. - M.: LLC "Agencija za medicinske informacije", 2005.
2. Pozdeev OK Medicinska mikrobiologija: udžbenik za med. sveučilišta. - M.: GEOTAR-MED, 2005. (monografija).
3. Korotyaev AI, Babichev SA Medicinska mikrobiologija, imunologija i virusologija / udžbenik za med. sveučilišta. - SPb.: SpetsLit, 2000 (monografija).
4. Vorobiev A.A., Bykov A.S., Paškov E.P., Rybakova A.M. Mikrobiologija: udžbenik. - M.: Medicina, 2003.
5. Medicinska mikrobiologija, virologija i imunologija: udžbenik / ur. V. V. Zvereva, M. N. Boychenko. - M.: GEOTar-Media, 2014 (priručnik).
6. Vodič za praktičnu nastavu iz medicinske mikrobiologije, virusologije i imunologije / ur. V.V. Teza. - M.: Medicina, 2002.
Sadržaj
Uvod 6
Sastav bakterija s gledišta njihove fiziologije. 7
Metabolizam 14
Prehrana (transport hranjivih tvari) 25
Visina 29
Dah 31
Uzgoj 34
Mikrobne zajednice 37
DODACI 49
Testovi 102
Literatura 105
…………..
…………
Neke ljudske bolesti povezane su s izlaganjem mikrobima na tijelu. Kada se takve bolesti pojave u ljudskom tijelu, dolazi do složenih promjena, mobiliziraju se zaštitne funkcije, usmjerene na borbu protiv zarobljenih mikroba.
Među ogromnim brojem mikroorganizama postoje oni koji su sposobni izazvati bolesti kod ljudi, životinja i biljaka. Nazivaju se patogenima ili uzročnicima bolesti. Specifičnost je karakteristična za patogene mikroorganizme - svaka vrsta njih može izazvati samo određenu bolest sa svim svojim karakterističnim znakovima. Većina patogenih mikroorganizama su parazitski mikrobi, budući da su sposobni živjeti od tvari živog organizma.
Patogeni mikrobi proizvode posebne tvari - toksine koji truju tijelo i uzrokuju bolno stanje. Sposobnost mikroorganizama da izazovu bolest naziva se patogenost. Može se manifestirati u različitim stupnjevima. Stupanj patogenosti naziva se virulencija.Virulencija mikroba može se povećati ili smanjiti u prirodnim i eksperimentalnim uvjetima.
Pod pojmom "infekcija»Shvatiti proces interakcije mikroba s ljudskim tijelom, uslijed čega dolazi do zarazne bolesti. Izvori infekcije su, prije svega, bolesni ljudi i životinje koji ispuštaju patogene u vanjsko okruženje, kao i bolesni ljudi i životinje u čijem se tijelu patogeni mikrobi nastavljaju zadržavati još neko vrijeme (ponekad vrlo dugo) nakon oporavka . Ljudi i životinje koji nakon oporavka oslobađaju patogene mikrobe nazivaju se prijenosnici bakterija ili izlučivači bakterija. Ljudi koji nisu bolesni također mogu biti prijenosnici bakterija. Patogeni mikrobi koje je izolirao bolesni organizam ulaze u zrak, tlo, vodu, okolne predmete i hranu, gdje mogu ostati održivi više ili manje dugo, ovisno o vrsti mikroba.
Patogeni mikrobi ulaze u ljudsko tijelo na različite načine: izravnim kontaktom s bolesnom osobom, zrakom s najmanjim kapljicama sluzi i sline koje pacijent izlučuje pri kašljanju ili kihanju (kapljična infekcija). Mikrobi - uzročnici nekih bolesti - izlučuju se bolesnici s izmetom i urinom. Takvi mikrobi ulaze u tijelo zdrave osobe kroz prljave ruke koje nisu oprane nakon korištenja toaleta. Zaražene ruke također šire patogene na hranu. Uzročnici crijevnih infekcija također prodiru u zdravo tijelo kada se konzumira zagađena voda.
Insekti i glodavci često su prijenosnici zaraznih bolesti. Muhe mogu prenositi uzročnike trbušnog tifusa i dizenterije, uši - tifus, neke vrste komaraca - malarije itd. Miševi i štakori distributeri su uzročnika kuge, tularemije, antraksa.
Prodiranje patogenih mikroba u ljudsko tijelo ne dovodi uvijek do njegove bolesti. U nastanku i tijeku bolesti važnu ulogu imaju svojstva patogenih mikroba koji su prodrli u tijelo, njihov broj i aktivnost, kao i mjesto njihovog unošenja u organizam i stanje samog organizma.
Za razmnožavanje patogenih mikroorganizama potrebni su povoljni uvjeti, a oni se pojavljuju kada je ljudsko tijelo oslabljeno i njegova zaštitna sposobnost smanjena (zbog hipotermije, gladovanja itd.). Osjetljivost na infekcije također ovisi o dobi (u djece i starijih osoba veća je nego u odraslih).
Od trenutka ulaska patogenih mikroba u ljudsko tijelo do pojave znakova bolesti obično prolazi određeno vrijeme - naziva se latentno razdoblje bolesti ili inkubacija. Za to vrijeme dolazi do umnožavanja patogenih mikroba u tijelu i nakupljanja štetnih produkata njihove vitalne aktivnosti. Trajanje razdoblja inkubacije za različite bolesti nije isto. U rasponu je od nekoliko dana (gripa, tetanus, dizenterija) do nekoliko tjedana (tifus, tifusna groznica), a ponekad dosegne i nekoliko mjeseci (bjesnoća) pa čak i godina (guba).
Ako pronađete pogrešku, odaberite dio teksta i pritisnite Ctrl + Enter.
Mediji za kulturu - podloge koje se sastoje od komponenti koje osiguravaju potrebne uvjete za uzgoj mikroorganizama ili nakupljanje njihovih otpadnih proizvoda
.
Mediji za kulturu razlikuju se po namjeni, konzistenciji i sastavu. Prema namjeni konvencionalno se dijele u dvije glavne skupine - dijagnostičko i proizvodno okruženje. Dijagnostički mediji za kulturu uključuju pet podskupina: mediji za uzgoj širokog spektra mikroorganizama; medij za izolaciju specifičnog patogena; različiti mediji koji omogućuju razlikovanje određenih vrsta mikroorganizama; medija za identifikaciju mikroorganizama i medija za pohranu namijenjenih obogaćivanju određene vrste mikroorganizama. Proizvodnja uključuje P. sa. koji se koriste u industrijskoj proizvodnji bioloških lijekova (bakterijska cjepiva, toksoidi itd.) i njihovu kontrolu kvalitete.Mediji za kulturu za proizvodnju bakterijskih pripravaka, za razliku od dijagnostičkih medija, ne smiju sadržavati nečistoće štetne za ljude i imati negativan učinak na proces proizvodnje (bez ometanja, osobito, uklanjanja produkata mikrobnog metabolizma, organskog balasta i mineralne tvari iz pripremljenih pripravaka).
Prema konzistenciji razlikuju se tekući, gusti i polutekući mediji. Čvrste i polutekuće hranjive podloge pripremaju se od tekućih podloga dodavanjem agara ili (rjeđe) želatine. Agar agar je polisaharid izveden iz određenih sorti morskih algi. Agar se obično uvodi u P. s. u koncentraciji 1-2%(u proizvodnji polutekućih medija-0,2-0,4%), želatine-10-15%. Na t ° 25-30 ° želatinozni medij se topi pa se na njima uzgajaju mikroorganizmi uglavnom na sobnoj temperaturi. Zgrušani krvni serum, valjana jaja, krumpir i mediji koji sadrže 1,5% silika gela također se koriste kao čvrsti podlozi.
Po sastavu mediji za kulturu dijele se na jednostavne i složene. Jednostavan P. sa. osiguravaju prehrambene potrebe većine patogenih mikroorganizama (mezopatamijska juha, mezopatamijski agar, Hottingerova juha i agar, hranjiva želatina, peptonska voda itd.). Složeni mediji uključuju posebne medije za mikrobe koji ne rastu na jednostavnim hranjivim podlogama. Takvi se mediji pripremaju dodavanjem krvi, seruma, ugljikohidrata i drugih tvari potrebnih za reprodukciju određenog mikroorganizma u jednostavne medije. Složeni P. s. su također diferencijalno dijagnostički mediji, na primjer, Gissovi mediji s ugljikohidratima i indikatorom, koji se koriste za određivanje vrste mikroba koji se proučava prema njegovoj enzimskoj aktivnosti. Neki složeni mediji, takozvani selektivni ili izborni, koriste se za ciljanu izolaciju jedne ili više vrsta mikroorganizama stvaranjem optimalnih uvjeta za njihov rast i inhibicijom rasta prateće mikroflore. Na primjer, bizmut sulfitni agar strogo je selektivan medij za izolaciju salmonele, agar i Levinov medij su slabo selektivni mediji za izolaciju enterobakterija.
Ovisno o sastavu početnih komponenti, razlikuju se i sintetski, polusintetski i prirodni mediji. Sintetički mediji čije su komponente točno poznate, au nešto manjoj mjeri polusintetičke, prikladne su i koriste se uglavnom za proučavanje fizioloških procesa mikroorganizama. Korištenje medija ove vrste omogućuje utvrđivanje minimalnih potreba pojedinih mikroorganizama za hranjivim tvarima i na temelju toga stvaraju hranjive podloge koje sadrže samo one spojeve koji su potrebni za rast datog mikroba. Prednost sintetičkih medija za kulturu je njihova standardizacija, međutim, upotreba tih medija je ograničena zbog visoke cijene i složenosti sastava mnogih od njih (često sadrže do 40 ili više sastojaka). Osim toga, osjetljivije su na neravnoteže između nekih P. komponenti. , osobito aminokiseline, a rast mikroorganizama na takvim medijima lakše se suzbija prekomjernim prozračivanjem ili otrovnim kationima.
U mikrobiološkoj praksi i dalje se naširoko koriste mediji prirodnog podrijetla čiji kemijski sastav nije dobro poznat. Temelj takvih medija sastoji se od različitih sirovina životinjskog ili biljnog podrijetla: mesa i njegovih nadomjestaka, ribe, životinjske krvi, kazeina, kvasca, krumpira, soje itd. Većim dijelom samog okoliša.
Uz proteinske baze, hranjivi mediji trebaju sadržavati elemente pepela (fosfor, sumpor, kalcij, magnezij, željezo) i elemente u tragovima (bor, molibden, kobalt, mangan, cink, nikal, bakar, klor, natrij, silicij itd.). Te su tvari potrebne za mnoge biosintetske procese u bakterijama, ali potreba za njima u različitih vrsta mikroorganizama nije ista. Na primjer, mangan i željezo potrebni su za patogene bakterije E. coli i kuge, a kalij za bakterije mliječne kiseline. Mangan, kalcij, magnezij, kalij i željezo potiču rast bacila antraksa, a magnezij, željezo i mangan potiču rast brucele.
Za uzgoj mnogih patogenih mikroorganizama nužna je prisutnost u mediju takozvanih faktora rasta, zastupljenih prvenstveno vitaminima topljivim u vodi. Iako bakterije ne koriste te tvari kao plastični ili energetski materijal, ipak su one neophodne komponente hranjivih medija, jer njihova odsutnost inhibira stvaranje mnogih enzima. Određene organske kiseline, purinske i pirimidinske baze i aminokiseline također mogu biti čimbenici rasta. Određene aminokiseline (L-cistin, D-piroglutaminska kiselina itd.) Mogu potaknuti rast nekih mikroorganizama i, obrnuto, inhibiraju rast drugih.
Mediji za kulturu mora sadržavati sve kemijske elemente potrebne za uzgojene mikroorganizme u lako usvojivom obliku i u optimalnim količinama. Izvor ugljika u P. s. obično se koriste odvojeni ugljikohidrati, soli organskih kiselina, kao i ugljik, koji je dio spojeva koji sadrže dušik (proteini, peptoni, aminokiseline itd.). Zahtjevi za dušikom ispunjeni su ako u medijima postoji izvorni životinjski protein (krv, serum, ascitna tekućina itd.), Peptoni, aminokiseline, amonijeve soli i druge tvari koje sadrže dušik (purinske i pirimidinske baze, urea itd.) . V. hranjivi mediji čine mineralne soli neophodne za mikroorganizme. Mikroelementi potrebni bakterijama u zanemarivim količinama obično ulaze u hranjive medije bilo s vodom, koja se koristi za pripremu podloge, ili sa sirovinama koje su dio hranjivog medija. Čimbenici rasta ulaze u okoliš s raznim dijalizatima, ekstraktima i autolizatima u obliku aminokiselina, peptida, purinskih i pirimidinskih baza i vitamina.
Sirovine koje sadrže bjelančevine koje se koriste za pripremu medija za kulturu hidroliziraju se pomoću različitih enzima (pepsin, tripsin, pankreatin, papain, gljivične proteaze itd.) Ili kiselina (rjeđe lužine). Svrha hidrolize je otapanje i razgradnja proteina uz stvaranje dušikovih spojeva koje asimilira mikrobna stanica: peptoni, polipeptidi, aminokiseline, koji su dio hidrolizata dobivenih na ovaj način. Da bi se zadovoljile prehrambene potrebe svake određene vrste mikroorganizama, koriste se ti ili oni hidrolizati, ovisno o njihovom kemijskom sastavu ili u sastavu P. s. istovremeno se uvodi nekoliko hidrolizata u korištenim omjerima.
Dizajnirano P. s. prema svom sastavu i fizikalno -kemijskim svojstvima mora odgovarati uvjetima prirodnog obitavanja mikroorganizama. Razlike u njihovim prehrambenim potrebama toliko su raznolike da isključuju mogućnost stvaranja univerzalnog P. s. Stoga se suvremena načela oblikovanja medija temelje na opsežnom proučavanju prehrambenih potreba mikroorganizama.
Potrebno je da hranjivi mediji ne sadrže samo hranjive tvari potrebne za mikrobe, već i da imaju optimalnu koncentraciju vodikovih i hidroksilnih iona. Većina patogenih mikroorganizama bolje raste na hranjivim medijima s blago alkalnom reakcijom (pH 7,2-7,4).Izuzetak su Vibrio cholerae čiji je optimalan rast u alkalnoj zoni (pH 8,5-9,0) i uzročnik tuberkuloze koji zahtijeva slabo kiselu reakciju (pH 6,2-6,8). Kako bi se spriječila promjena pH tijekom uzgoja mikroorganizama u P. sa. dodati fosfatne pufere - mješavinu mono- i disupstituiranih kalijevih fosfata čija koncentracija u mediju ne smije prelaziti 0,5%.
Mediji za kulturu mora imati dovoljnu vlažnost i biti izotoničan za mikrobnu stanicu, što osigurava normalan tijek najvažnijih fizikalnih i kemijskih procesa u njoj. Za većinu mikroorganizama optimalno okruženje je odgovarajuća 0,5% -tna otopina natrijevog klorida. Jedan od bitnih zahtjeva za kulturne medije je njihova sterilnost, što omogućuje uzgoj čistih kultura mikroba.
Važnu ulogu u P. -ovom prijemu po stranici. odgovarajuća kvaliteta pripada metodama njihove kontrole. Za hranjive podloge koje se koriste u proizvodnji, glavni pokazatelj kvalitete je učinkovitost medija (prinos), tj. sposobnost akumuliranja najveće količine biomase mikroorganizama punih svojstava ili produkata njihove biosinteze (toksini itd.). U procjeni dijagnostičkih hranjivih medija vodeći kriterij je pokazatelj osjetljivosti - sposobnost osiguravanja rasta mikroorganizama u maksimalnim razrjeđenjima kulture patogena. Ovisno o namjeni hranjivih podloga, pri procjeni njihove kakvoće koriste se i drugi pokazatelji - stabilnost glavnih svojstava uzgojenih mikroorganizama i brzina njihovog rasta, ozbiljnost svojstava razlikovanja itd.
Pri rješavanju pitanja kvalitete hranjivih medija važna važnost pridaje se njihovoj standardizaciji, što olakšava proizvodnja medija u obliku suhih pripravaka. U SSSR -u je uspostavljena industrijska proizvodnja suhih medija za različite namjene: jednostavne, selektivne, diferencijalno dijagnostičke i posebne.
Bibliografija: Yu.A. Kozlov Mediji za kulturu u medicinskoj mikrobiologiji, M., 1950, bibliogr.; Laboratorijske metode istraživanja u klinici, ur. V.V. Menšikov, s. 315, 343, M., 1987 .; Meinell J. i Meinell E. Eksperimentalna mikrobiologija, prev. s engleskog, str. 46, M., 1967 .; Mikrobiološke metode istraživanja zaraznih bolesti, ur. G.Ya. Sinaj i O.G. Birger, s. 64, M., 1949; Enterobacteriaceae, ur. U I. Pokrovsky, str. 258, M., 1985. (zbornik).
Pažnja! Članak 'Mediji za kulturu'Pruža se samo u informativne svrhe i ne smije se koristiti za samoliječenje


