Titlovi
Licensingedit
Ja, nositelj autorskih prava nad ovim djelom, ovim ga objavljujem pod sljedećom licencom:
|
Povijest datoteka
Kliknite na datum / vrijeme da biste vidjeli datoteku u tom trenutku.
| Datum vrijeme | Minijatura | Dimenzije | Korisnik | Komentar | |
|---|---|---|---|---|---|
| Trenutno | 07:28, 5. kolovoza 2014. godine | 3.022 × 2.341 (1,19 MB) | Selso | Stranica koju je izradio korisnik pomoću UploadWizarda |
Sljedeća stranica koristi ovu datoteku:
Datoteka: Thelephora anthocephala a1 (4) .JPG (preusmjeravanje datoteke)
Sljedeće druge wiki -je koriste ovu datoteku:
- Korištenje na pl.wikipedia.org
Metapodaci
Ova datoteka sadrži dodatne informacije, poput Exif metapodataka koje je možda dodala digitalna kamera, skener ili softver koji se koristi za njihovu izradu ili digitalizaciju. Ako je datoteka izmijenjena u izvornom stanju, neki detalji, poput vremenske oznake, možda neće u potpunosti odražavati podatke izvorne datoteke. Vremenska oznaka točna je samo koliko i sat u kameri, a može biti i potpuno pogrešna.
| Proizvođač fotoaparata | SONY |
|---|---|
| Model kamere | DSC-HX100V |
| Vrijeme izloženosti | 1/100 s (0,01) |
| F-broj | f / 2,8 |
| ISO ocjena brzine | 100 |
| Datum i vrijeme generiranja podataka | 00:06, 4. kolovoza 2014. godine |
| Žarišna duljina objektiva | 4,8 mm |
| Orijentacija | Normalan |
| Vodoravna rezolucija | 72 dpi |
| Okomita rezolucija | 72 dpi |
| Datum i vrijeme promjene datoteke | 00:06, 4. kolovoza 2014. godine |
| Y i C pozicioniranje | Su-smješteno |
| Program izlaganja | Normalan program |
| Exif verzija | 2.3 |
| Datum i vrijeme digitalizacije | 00:06, 4. kolovoza 2014. godine |
| Značenje svake komponente |
|
| Način kompresije slike | 1 |
| APEX svjetlina | 4.76328125 |
| Pristranost izloženosti vrhovima | |
| Maksimalni otvor zemlje | 3 APEX (f / 2,83) |
| Način mjerenja | Uzorak |
| Izvor svjetlosti | Nepoznato |
| Bljesak | Blic se nije aktivirao, automatski način rada |
| Podržana verzija Flashpixa | 0,100 |
| Prostor u boji | sRGB |
| Izvor datoteke | Digitalni fotoaparat |
| Vrsta scene | Izravno fotografirana slika |
| Prilagođena obrada slike | Normalan proces |
| Način ekspozicije | Automatska ekspozicija |
| balans bijele boje | Automatski balans bijele boje |
| Omjer digitalnog zuma | 1 |
| Vrsta snimanja scene | Standard |
| Kontrast | Normalan |
| Zasićenje | Normalan |
| Oštrina | Normalan |
Opis
Voćna tijela tipično su hrskavičasta ili gumasto-želatinozna. U izlivenim vrstama (onima koje su rahlo ravne ili ravne) formiraju se na površini tla ili u steljni listići, često inkrustrirajući pale grančice i krhotine, ponekad inkrustrirajući osnove stabljika živih biljaka. U tipičnih vrsta mogu se također pojaviti nepravilni izdanci ili koralji. U jednoj vrsti nastaju izdanci slični zagradama. Kod dvije druge vrste plodovi su u potpunosti nalik koraljima ili mreži. Spore su bijele mase.
Mikroskopski likovi
Voćna tijela sastavljena su od hifa kojima nedostaju stezaljke u želatinoznoj matrici. U jednoj vrsti hifalni sustav je dimitičan. Površina koja nosi spore u početku je prekrivena slojem slabo razgranate hifidije ispod koje nastaju bazidije. Bazidije su tremeloidne (elipsoidne i okomito septirane), stvarajući duge, vijugave sterigmate ili epibasidije na kojima se stvaraju bazidiospore. Te su spore tipično elipsoidne do duguljaste, ali u jednoj vrsti alantoidne (u obliku kobasice), a u druge crvolike (u obliku crva).
Taksonomija
Rod su prvi put objavili 1871. godine Louis i Charles Tulasne koji su otkrili da dvije vrste (Sebacina incrustans i S. epigaea) prethodno spomenuto Korticij ili Telefora posjedovali su septirane bazidije, slične onima koje se nalaze u rodu Tremella... Iako je u to vrijeme bilo neobično razdvajati rodove gljiva na čisto mikroskopskim obilježjima, Sebacina podignuta je za ispuštanje, Korticij-like gljive s tremeloidnom bazidijom.
Naknadni su autori rodu dodali mnoge dodatne vrste. Većina se, međutim, pokazala nepovezanom Sebacina a uklonio ih je 1957. Ervin, koji je promiješao neke vrste u Heterochaetella (na primjer, Heterochaetella dubia, sada Stypella dubia (Bourdot & Galzin) P. Roberts), Bourdotia i Exidiopsis... Godine 1961. Wells je također prenio mnoge vrste u rod Exidiopsis... Tip vrsta, S. incrustans, ima promjenjivu morfologiju, što je dovelo do toga da mu se dodeli niz imena.
Nedavna molekularna istraživanja su to pokazala Sebacina daleko je raznolikiji nego što se ranije pretpostavljalo, iako se ta genetska raznolikost ne mora odražavati u morfološkim obilježjima (drugim riječima, vrste je nemoguće razlikovati osim analizom DNA). Isto istraživanje također pokazuje da se rod dijeli u dvije skupine, jedna grupirana s tipičnom vrstom, druga skupljena s vrstom koja se ponekad naziva i "Sebacina vermifera" (iako je to pogrešna identifikacija). Ova posljednja vrsta razlikuje se po formiranju tankih, voštanih plodova, a skupina možda nije dio Sebacina u strogom smislu. Čak i u strogom smislu, Sebacina je umjetno i ne razlikuje se jasno od srodnih rodova poput koralja Tremellodendron.
taksonomija
El género fue publicado por primera vez en 1871 por Louis i Charles Tulasne que había descubierto que dos especies ( inkrustans Sebacina y S. Epigaea ) antes referidos Korticij o Telefora poseían septadas basidios, similares a los encontrados en el género Tremella ... Pesar de que era inusual en ese momento para separar géneros de hongos en los personajes puramente microscópicas, Sebacina fue erigido para effused, Korticij -como hongos con basidios tremelloid.
Autores posteriores añadieron muchas especies adicionales al género. La Mayoría, embargo na grijeh, rezultató no relacionado con Sebacina y fueron retirados en 1957. por Ervin, remató de algunas especies a Heterohetela (na primjer, Heterochaetella dubia , ahora Stypella dubia (Bourdot y Galzin) P. Roberts), Bourdotia y Exidiopsis ... Godine 1961., Wells también transfirió a muchas especies del género Exidiopsis ... Los especie tipo, S. incrustans , tiene una variable de la morfología, lo que ha llevado a que se le asigna un número de nombres.
Recientes investiciones molecular ha demostrado que Sebacina es mucho más diverso que asumido previamente, aunque esta diversidad genética puede no quedar reflejada en los caracteres morfológicos (en otras palabras, las especies pueden ser imposibles de distirir excepto por análisis de ADN). La misma investición también muestra que el género se divide en dos grupos, uno agrupado con la especie tipo, el otro agrupado con una especie refiere a veces como “Sebacina vermifera” (aunque esto es un error de identificación). Esta última especie es distita en la formación de cuerpos frutales delgadas, cerosos y el grupo puede no ser parte de Sebacina en el sentido estricto. Uključujući en el sentido estricto, Sebacina es umjetno, no siendo claramente distintas de géneros relacionados, tales como el coral-como Tremellodendron .










Definitor
- Basidia
-
Lat. Basidia. Specijalizirana struktura spolnog razmnožavanja u gljivama, svojstvena samo basidiomicetima. Basidije su završni (krajnji) elementi hifa različitih oblika i veličina, na kojima se spore egzogeno razvijaju (izvana).
Basidije su različite po strukturi i načinu vezivanja za hife.
Prema položaju u odnosu na os hife, na koju su pričvršćene, razlikuju se tri vrste bazidija:
Apikalne bazidije nastaju od terminalne stanice hife i nalaze se paralelno s osi.
Pleurobasidije nastaju iz lateralnih procesa i nalaze se okomito na os hife, koja nastavlja rasti i može stvarati nove procese s bazidijama.
Subasidije nastaju iz lateralnog procesa, okrenutog okomito na os hife, koja nakon formiranja jednog bazidija zaustavlja njegov rast.
Na temelju morfologije:
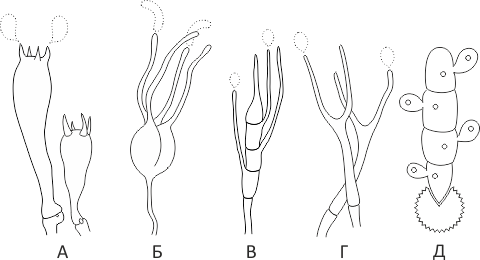
Holobasidia - jednostanične bazidije, nisu podijeljene pregradama (vidi sliku A, D.).
Fragmobasidije su podijeljene poprečnim ili okomitim pregradama, obično u četiri stanice (vidi sliku B, C).
Prema vrsti razvoja:
Heterobasidija se sastoji od dva dijela - hipobasidije i epibasidije koja se razvija iz nje, sa ili bez pregrada (vidi sliku C, B) (vidi sliku D).
Homobasidije se ne dijele na hipo- i epibasidije te se u svim slučajevima smatraju holobasidijama (slika A).
Basidia je mjesto kariogamije, mejoze i stvaranja basidiospora. Homobasidija u pravilu nije funkcionalno podijeljena, a mejoza u njoj slijedi kariogamiju. No, bazidije se mogu podijeliti na probasidije - mjesto kariogamije i metabasidije - mjesto mejoze. Probasidija je često uspavana spora, na primjer u gljivama hrđe. U takvim slučajevima probazidija raste s metabazidijama, u kojima dolazi do mejoze i na kojima nastaju bazidiospore (vidi sliku E).
Vidi Kariogamija, Mejoza, Gifa.
- Basidia (Basidia)
-
Lat. Basidia. Specijalizirana struktura spolnog razmnožavanja u gljivama, svojstvena samo basidiomicetima. Basidije su završni (krajnji) elementi hifa različitih oblika i veličina, na kojima se spore egzogeno razvijaju (izvana).
Basidije su različite po strukturi i načinu vezivanja za hife.
Prema položaju u odnosu na os hife, na koju su pričvršćene, razlikuju se tri vrste bazidija:
Apikalne bazidije nastaju od terminalne stanice hife i nalaze se paralelno s osi.
Pleurobasidije nastaju iz lateralnih procesa i nalaze se okomito na os hife, koja nastavlja rasti i može stvarati nove procese s bazidijama.
Subasidije nastaju iz lateralnog procesa, okrenutog okomito na os hife, koja nakon formiranja jednog bazidija zaustavlja njegov rast.
Na temelju morfologije:
Holobasidia - jednostanične bazidije, nisu podijeljene pregradama (vidi sliku A, D.).
Fragmobasidije su podijeljene poprečnim ili okomitim pregradama, obično u četiri stanice (vidi sliku B, C).
Prema vrsti razvoja:
Heterobasidija se sastoji od dva dijela - hipobasidije i epibasidije koja se razvija iz nje, sa ili bez pregrada (vidi sliku C, B) (vidi sliku D).
Homobasidije se ne dijele na hipo- i epibasidije te se u svim slučajevima smatraju holobasidijama (slika A).
Basidia je mjesto kariogamije, mejoze i stvaranja basidiospora. Homobasidija u pravilu nije funkcionalno podijeljena, a mejoza u njoj slijedi kariogamiju. No, bazidije se mogu podijeliti na probasidije - mjesto kariogamije i metabasidije - mjesto mejoze. Probasidija je često uspavana spora, na primjer u gljivama hrđe. U takvim slučajevima probazidija raste s metabazidijama, u kojima dolazi do mejoze i na kojima nastaju bazidiospore (vidi sliku E).
Vidi Kariogamija, Mejoza, Gifa.
-










Opis
cuerpos de frutas son típicamente cartilaginoso o de caucho-gelatinosa. En las especies effused (aquellas que se extienden sin apretar o planu), que se forman sobre la superficie del suelo o en la hojarasca, a menudo incrustantes ramas caidos y escombros, a veces incrustantes las bases de los tallos de las plantas vivas. En la especie tipo, excrecencias unregulares o de coral-como también se pueden producir. En una especie, se forman excrecencias soporte similar. En otras dos especies, los cuerpos fructíferos son totalmente coral- o en forma de red. Las esporas son blancas en masa.
caracteres microscópicos
Cuerpos de frutas se komponen de hifas que carecen de conexiones de abrazadera en una matriz želatinoza. En una de las especies del sistema de las hifas es dimitic. La espora superficie -bearing está cubierto inicialmente en una capa de débilmente ramificados hyphidia por debajo del cual los basidios se forman. Los basidios se tremelloid (elipsoide y septadas verticalmente), dando lugar a, largo y sinuoso esterigmas o epibasidia en que las basidiosporas se producen. Estas esporas son típicamente elipsoide a oblongo, pero alantoides (salchicha con forma) en una especie y vermiforme (forma de gusano) en otro.










Taksonomija
Rod su prvi put objavili 1871. godine Louis i Charles Tulays, koji su otkrili da dvije vrste (Sebacina incrustans i S. epigaea) prethodno spomenuto Korticij, ili Telefora posjedovali su septirane bazidije slične onima koji se nalaze u rodu Tremella... Iako je u to vrijeme bilo neobično razlikovati rodove gljiva na čisto mikroskopskim obilježjima, Sebacina postavljeno je za izlivanje, poput Corticiuma gljive s tremeloidnom bazidijom.
Kasniji su autori u rod dodali mnoge dodatne vrste. Međutim, pokazalo se da većina nije povezana Sebacina a 1957. uklonio ga je Erwin, koji je promiješao neke vrste u Heterohetela (na primjer, Heterochaetella dubia, sada Stypella dubia (Bourdot & Galzin) P. Roberts), Bourdotia i Exidiopsis... Wells je također prenio mnoge vrste na rod 1961. godine. Exidiopsis... Različite vrste, S. incrustansPostoji promjenjiva morfologija koja je dovela do toga da se dodijeli mnogim imenima.
Nedavna molekularna istraživanja su to pokazala Sebacina mnogo je raznolikiji nego što je prije prihvaćeno, iako se ta genetska raznolikost ne može odraziti u morfološkim znakovima (drugim riječima, osim vrsta, možda se neće moći razlikovati analizom DNA). Ista studija također pokazuje da se rod dijeli u dvije skupine, jedna grupirana s tipičnim vrstama, druga s vrstom koja se ponekad naziva "Sebacina vermifera" (iako je to pogrešna identifikacija). Ova druga sorta razlikuje se po stvaranju tankih, voštanih voćnih tijela, pa skupina možda nije dio Sebacina u strogom smislu. Čak i u strogom smislu, Sebacina umjetno, a da se ne razlikuje jasno od srodnih rodova, poput koralja Tremellodendron.
Vrste
sparassoidea Sebacina
Para los nombres de más edad, la lista se basa en especies mantenidas en el Sebacina por Wells (1961.).
- Sebacina allantoidea - descrito de Alemania
- Sebacina alutacea - descrito de la India)
- Concrescens Sebacina - descritos de los EE.UU.
- Sebacina dimitica - descrito de Alemania
- Sebacina Epigaea - descrito de Inglaterra
- Sebacina guayanensis - Venezuela
- Helvelloides Sebacina - descritos de los EE.UU.
- Incrustans Sebacina - descritos de Europa
- Sebacina pileata - descrito de Belice
- Sebacina sparassoidea - descrito de los EE.UU.
- Sebacina esponjosa - descrito de las Bahami
- Sebacina tomentosa - Venezuela
Osnovne informacije
iskaznica
533386049
Možete uređivati:
Ne
Mogu se sakriti postavkama privatnosti:
Ne
Jedinstveni identifikator korisnika, utvrđen tijekom registracije.
Domena
id533386049
Možete uređivati:
Da
Potrebno popuniti:
Ne
Mogu se sakriti postavkama privatnosti:
Ne
Domena se koristi za postavljanje lijepe, nezaboravne veze na stranicu korisnika.
Ime
Elle
Možete uređivati:
Da
Potrebno popuniti:
Da
Mogu se sakriti postavkama privatnosti:
Ne
Prezime
Sebacina
Možete uređivati:
Da
Potrebno popuniti:
Da
Mogu se sakriti postavkama privatnosti:
Ne
srednje ime
nije naznačeno
Možete uređivati:
Ne
Potrebno popuniti:
Ne
Mogu se sakriti postavkama privatnosti:
Ne
više nije moguće uređivati srednje ime za korisnike koji ga prethodno nisu naveli.
Kat
muški
Možete uređivati:
Da
Potrebno popuniti:
Da
Mogu se sakriti postavkama privatnosti:
Ne
Datum rođenja
27. ožujka 2003
Možete uređivati:
Da
Potrebno popuniti:
Da
Mogu se sakriti postavkama privatnosti:
Da
postoji mogućnost da se u cijelosti ili djelomično sakrije datum rođenja (u ovom slučaju bit će prikazani samo dan i mjesec rođenja).
Opis
Voćna tijela tipično su hrskavična ili elastično-želatinozna. U izlivenim vrstama (onima koje se slobodno šire ili ravne) formiraju se na površini tla ili u krhotinama lišća, često inkrustrirajući otpale grane i krhotine, a ponekad i temeljno korijenje živih biljaka. U tipičnih vrsta mogu se proizvoditi i nepravilna hrana ili hrana poput koralja. U jednoj vrsti nastaju namirnice slične zagradama. U druge dvije vrste plodna su tijela u potpunosti koraljna ili u obliku rešetki. Spore su bijele mase.






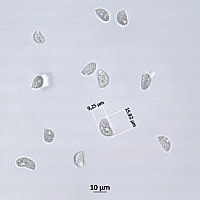



Mikroskopske oznake
Voćna tijela sastavljena su od hifa kojima nedostaju prstohvatne veze u želatinoznoj matrici. U jednoj vrsti hifalni sustav je dimitičan. Površina spora u početku je prekrivena slojem slabo razgranate hifidije, ispod koje nastaju bazidije. Bazidije su tremeloidne (elipsoidne i uspravne pregrade), nastaju duge, vijugave sterigmate ili epibasidije na kojima se stvaraju bazidiospore. Ove su spore tipično elipsoidne do duguljaste, ali (u obliku kobasice) u jednoj sorti, a (u obliku crva) u drugoj.
Metapodaci
Ova datoteka sadrži dodatne informacije, poput Exif metapodataka koje je možda dodala digitalna kamera, skener ili softver koji se koristi za njihovu izradu ili digitalizaciju. Ako je datoteka izmijenjena u izvornom stanju, neki detalji, poput vremenske oznake, možda neće u potpunosti odražavati podatke izvorne datoteke. Vremenska oznaka točna je samo koliko i sat u kameri, a može biti i potpuno pogrešna.
| Proizvođač fotoaparata | SONY |
|---|---|
| Model kamere | DSC-HX100V |
| Vrijeme izloženosti | 1/100 s (0,01) |
| F-broj | f / 2,8 |
| ISO ocjena brzine | 100 |
| Datum i vrijeme generiranja podataka | 00:06, 4. kolovoza 2014. godine |
| Žarišna duljina objektiva | 4,8 mm |
| Orijentacija | Normalan |
| Vodoravna rezolucija | 72 dpi |
| Okomita rezolucija | 72 dpi |
| Datum i vrijeme promjene datoteke | 00:06, 4. kolovoza 2014. godine |
| Y i C pozicioniranje | Su-smješteno |
| Program izlaganja | Normalan program |
| Exif verzija | 2.3 |
| Datum i vrijeme digitalizacije | 00:06, 4. kolovoza 2014. godine |
| Značenje svake komponente |
|
| Način kompresije slike | 1 |
| APEX svjetlina | 4.76328125 |
| Pristranost izloženosti vrhovima | |
| Maksimalni otvor zemlje | 3 APEX (f / 2,83) |
| Način mjerenja | Uzorak |
| Izvor svjetlosti | Nepoznato |
| Bljesak | Blic se nije aktivirao, automatski način rada |
| Podržana verzija Flashpixa | 0,100 |
| Prostor u boji | sRGB |
| Izvor datoteke | Digitalni fotoaparat |
| Vrsta scene | Izravno fotografirana slika |
| Prilagođena obrada slike | Normalan proces |
| Način ekspozicije | Automatska ekspozicija |
| balans bijele boje | Automatski balans bijele boje |
| Omjer digitalnog zuma | 1 |
| Vrsta snimanja scene | Standard |
| Kontrast | Normalan |
| Zasićenje | Normalan |
| Oštrina | Normalan |
Vrsta
 Sebacina sparassoidea
Sebacina sparassoidea
Za starija imena, popis se temelji na vrstama koje se zadržavaju u Sebacina Wells (1961).
- Sebacina allantoidea - opisano iz Njemačke
- Sebacina alutacea - opisano iz Indije)
- Sebacina concrescens - opisano iz SAD -a
- Sebacina dimitica - opisano iz Njemačke
- Sebacina epigaea - opisano iz Engleske
- Sebacina guayanensis - Venezuela
- Sebacina helvelloides - opisano iz SAD -a
- Sebacina incrustans - opisano iz Europe
- Sebacina pileata - opisano je iz Belizea
- Sebacina sparassoidea - opisano iz SAD -a
- Sebacina spongiosa - opisano s Bahama
- Sebacina tomentosa - Venezuela